Chapter: Psychology: Sensation
Vision Perceiving Shapes

Perceiving
Shapes
The perception of color enhances
our appreciation of art and, more practically, allows us to distinguish a ripe
fruit from a green one. Other aspects of vision are far more important. After
all, a color-blind individual can live a perfectly normal life. But the same
can’t be said for an individual who can’t tell a square from a circle, and
can’t tell whether the shape in front of her is that of an apple or that of a
banana. These individuals (known as visual
agnosics) are dramatically impaired in their functioning. We therefore need
to ask how the visual system manages the perception of shape.
FEATURE DETECTORS
Recordings from individual nerve
cells have allowed electrophysiologists to examine how particular cells in the
visual system respond to certain stimuli. In these studies, researchers place a
microelectrode into the optic nerve—or, in many studies, into the brain of an
anesthetized animal. The animal’s well-being is carefully monitored, both for
ethical reasons and to allow the investigators to assess how neurons function
in an intact, healthy organism. The animal’s eye is then stimulated by visual
inputs of varying brightness and different shapes, arriving at the eye from
different locations (Figure 4.41). In this way, the investigator can learn
which stimuli evoke a response from that cell.
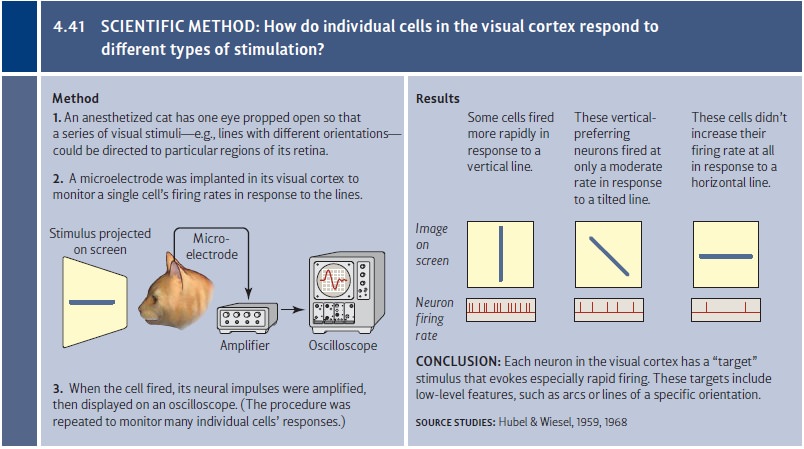
Results from these studies show
that the cells in the visual system—whether we’re considering the rods and
cones themselves, neurons in the optic nerve, or neurons in the brain—all have
a preferred target, a certain type of stimulus that’s especially effec-tive in
causing that cell to fire. We can think of the cells, therefore, as
“detectors,” each one tuned for (and so likely to detect) its own set of
targets.
What sorts of detectors does the
visual system rely on? The answer depends on the species. Frogs, for example,
need only a few bits of information about the world: “What’s that large shape
moving toward me? Just in case it’s a predator, I’ll take a leap to safety.”
“What’s that small, dark shape moving around? It might be a fly, so I think
I’ll flick my tongue at it.” Because they need so little information to
survive, frogs have just a few detector types—and they’re located on the
retina, so the frog can quickly ana-lyze the input and act on it (Lettvin,
Maturan, McCulloch, & Pitts, 1959).
Unlike frogs, most
animals—including the mammals—need more detailed infor-mation about the world
around them. Their visual systems perform a more complex analysis, supported by
a greater variety of detector types, located in the cortex as well as on the
retina. Most of what we know about this visual analysis comes from the work

of David Hubel and Torsten
Wiesel, who won a Nobel Prize for their research (Figure 4.42). Working first
with cats and then with primates, these investigators confirmed that each cell
in the visual cortex responds to stimuli in only a limited region of space—or,
equivalently, each cell in the retina responds to stimuli on only a limited
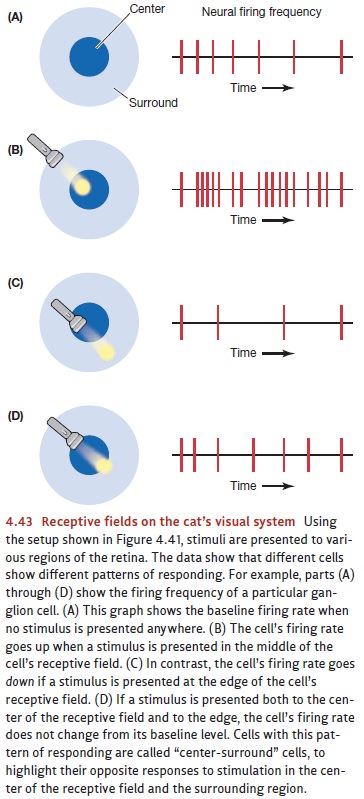
region of the retina. This region defines that cell’s receptive field (Figure 4.43). More important, this research made
it plain that cells differ in the types of detectors they are. Some cells have
receptive fields of a special size, location, and shape—and so they fire at their maximum rate only when the
visual input is a line of a specific orientation at a specific retinal
position. One such cell might respond to a vertical line at one posi-tion in
the visual field, while another cell might respond to a line tilted to 45
degrees at the same position; still another cell might respond to a vertical
line at some other position. In this way, and because the visual field is
blanketed by receptive fields, lines of any orientation at any position will be
detected by the appropriate type of cell (Hubel & Wiesel, 1959, 1968).
Other cells in the visual cortex
are a bit more sophisticated. They also fire only in response to a line or edge
of a particular orientation, but they’re largely indifferent to the line’s
specific location within the visual field (see Figure 4.41). Cells like these
serve as feature detectors,
detecting certain elements within the visual pattern. Other cells, deeper
within the visual system, presumably then assemble these now detected elements
in order to detect larger configurations and more complex patterns.
Consistent with this suggestion, Hubel and Wiesel were able to locate other cells that responded only to more complicated inputs. For example, some cells responded maximally to corners or particular angles. Other cells responded to movement pat-terns, firing maximally only when they detected movement of the appropriate velocity and direction.
DETECTORS FOR COMPLEXFORMS
It’s easy to see how feature
detectors might be useful. Let’s say that a pattern of visual input reaches
your retina and triggers a “horizontal-line detector” and a “vertical-line
detector.” When these two detectors fire at the same time, this event might
trigger a response in a “right-angle detector.” If at the same time you’ve also
detected three other right angles, this combination might trigger a response
from your “square detector.” Activity in this detector might then combine with
activity in other detectors and even-tually lead you to say, “Oh, look—there’s
my Intro Psych textbook.”
In this way, the feature
detectors we’ve just discussed might be the starting point for a hierarchy of
detectors that leads, step-by-step, from simple features to slightly more
complex shapes to still more complex shapes. And eventually, the steps will
lead to detectors that fire only in response to the sight of a complex object
in the world—such as the letter Q, or
your mother’s face, or a sports car.
Is this explanation plausible?
Could we have specific detectors for each of the count-less things we
recognize? For some targets, these detectors do exist—and, no surprise, they’re
usually detectors for stimuli that are especially significant for a species.
For example, certain cells in a monkey’s cortex have been shown to respond to
pictures of a monkey’s face, but not at all to pictures of other parts of a
monkey’s body. Other cells seem to respond almost exclusively to pictures of a
monkey’s hand—whether the hand has an open palm or clenched fist or the fingers
are pointed up or down (Desimone, Albright, Gross, & Bruce, 1984).
Still, it seems highly unlikely
that such built-in mechanisms could account for all the forms that higher
animals—especially humans—perceive and recognize. Simple crea-tures like frogs
are able to recognize only a few patterns, so it’s reasonable to think they
might have specialized detectors for each one. In contrast, humans easily
discriminate among a multitude of patterns; and this simple fact speaks
powerfully against the idea that we might have specialized detectors for each
of them—triangles, squares, apples, apple pies, champagne bottles, cabbages,
kings—the list is endless. We know that the perception of any kind of form
begins with a feature analysis; this process is plain in the functioning of the
detector cells. But how do we integrate these features to create more complex
forms? The answer to this question is surprisingly complex.
Related Topics