Chapter: Psychology: Sensation
The Visual Receptors
The Visual
Receptors
Once light reaches the retina, we
leave the domain of optics and enter that of neuro-physiology, because it is at
the retina that the physical stimulus energy is transduced into a neural
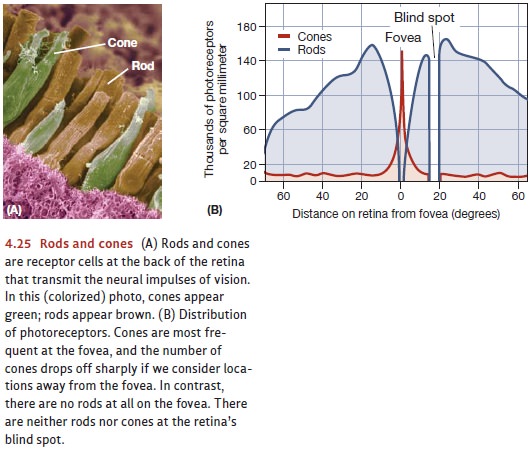
impulse. The retina contains two kinds of receptor cells, the rods and the cones; the names of these cells reflect their different shapes
(Figure 4.25). The cones areplentiful in the fovea, a small, roughly circular region at the center of the
retina; but they become less and less prevalent at the outer edges of the
retina. The opposite is true of the rods; they’re completely absent from the
fovea but more numerous at the retina’s edges. In all, there are some 120
million rods and about 6 million cones in the normal human eye.
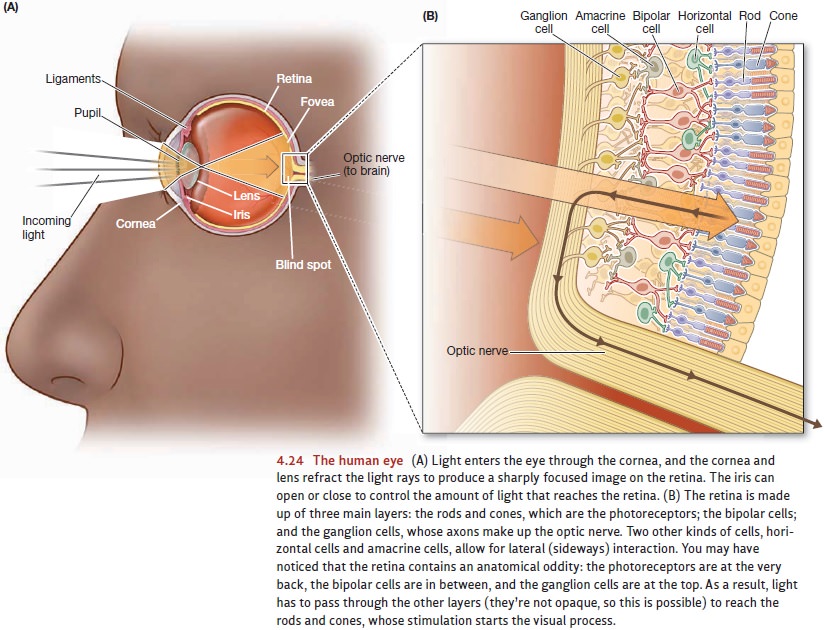
The rods and cones do not report
to the brain directly. Instead, their mes-sage is relayed by several other
layers of cells within the retina (see Figure 4.24). The receptors stimulate
the bipolar cells, and these in turn
excite the ganglioncells. The
ganglion cells collect informa- tion from all over the retina, and the axons of
these cells then converge to form a bundle of fibers that we call the opticnerve. Leaving the eyeball, the
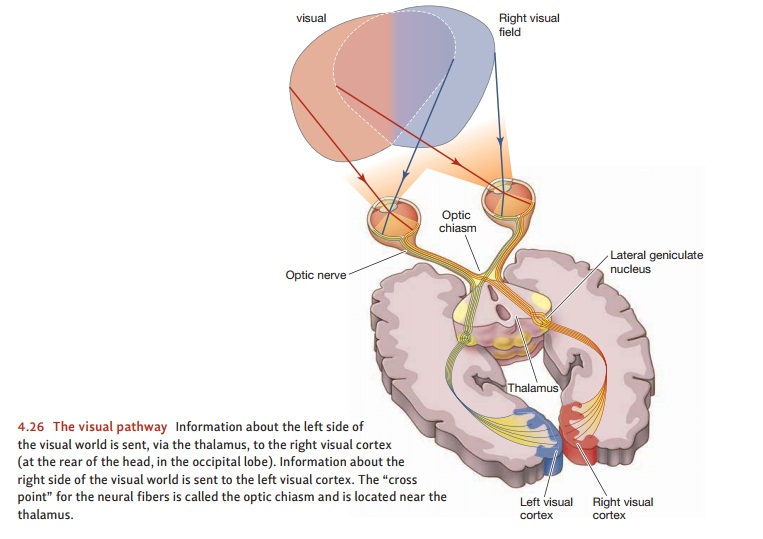
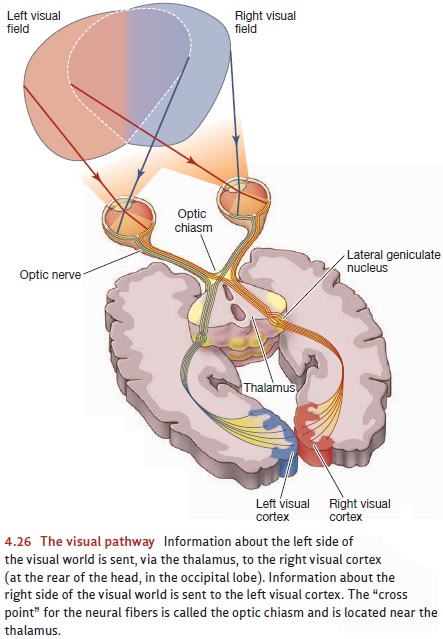
optic nerve car- ries information first to the lateral geniculatenucleus in the thalamus and then to the cor-tex
(Figure 4.26). (Notice that this pathway resembles the one for audi-tory
signals, which go from the ear to a different section of the geniculate nucleus
and then to the cortex.)
This anatomical arrangement
requires a space at the back of each eyeball to enable the axons of the
ganglion cells to exit the eye on their way to the thalamus. These axons fill
this space entirely, leaving no room for rods or cones. As a result, this region
has no photoreceptors and is completely insensitive to light. Appropriately
enough, it’s called the blind spot
(Figure 4.27).

Rods and cones differ in their
structure, number, and placement on the retina; they also differ in their
function. The rods are the receptors for night vision; they operate at low
light intensities and lead to achromatic
(colorless) sensations. The cones serve day vision; they respond at much higher
levels of illumination and are responsible for sensations of color.
Why do we need two types of
photoreceptors? The answer is clear when we consider the enormous range of
light intensities encountered by organisms like ourselves as we go about our
business during both day and night. In humans, the ratio in energy level
between the dimmest stimulus we can detect and the brightest we can tolerate is
roughly 1:100,000,000,000. Natural selection has allowed for this incredible
range by a biological division of labor—so we have two separate receptor
systems, one for vision in dim light and the other for vision in bright light.
The enormous sensitivity of the
rods comes at a price: The same traits that make the rods sensitive to low
levels of light also make them less able to discriminate fine detail. As a
result, acuity—the ability to
perceive detail—is much greater in the cones. This is the major reason why we
point our eyes toward any target that we’d like to perceive in detail. This
action positions our eyes so that the image of the target falls onto the fovea,
where the cones are most closely packed and visual acuity is greatest.
Be aware that the differences
between rods and cones also create situations in which we want to rely on the
rods. That’s why it’s sometimes helpful to look at something “out of the
corner” of your eye. Sailors and astronomers have known for years that when
you’re trying to find a barely visible star, it’s best not to look directly at
the star’s loca-tion. By looking slightly away from the star, you can ensure
that the star’s image falls outside of the fovea and onto a region of the
retina that’s dense with the more light-sensitive rods. This strategy limits
the ability to discern detail; but, by relying on the rods, it maximizes visual
sensitivity to faint stimuli.
Rods and cones can also be
distinguished in one further way—their chemistry. Inside each photoreceptor is
a photopigment, a light-sensitive
chemical pigment that allows the transduction of light energy into a neural
signal. When light enters the receptor, the light energy changes the chemical
form of the photopigment, setting off a chain of events that ultimately leads
to an electrical signal. In this way, the light energy is translated into the
electrochemical language of the nervous system. Inside the receptor, the
pigment itself is then reconstituted so that it will be ready to react with
light again when the next opportunity arises.
Rods and cones contain different photopigments. The rods contain rhodopsin, a pig-ment that breaks down more readily in response to light than the cone pigments do.
Rhodopsin is part of the reason
that rods can function at lower light levels. There are three different cone
photopigments, and each cone contains one of the three types. The differ-ences
among the three pigments are crucial to the cones’ ability to discriminate
colors—a topic we’ll turn to shortly. Rods, which contain just one pigment, are
sensitive to differ-ences in brightness (white versus gray, or a strongly
illuminated red versus a weakly illuminated one); but they cannot discriminate
among different hues. So, for example, the rods will respond in exactly the
same way to a patch of red and an equally bright patch of blue. In effect, this
response makes each of us nearly “color blind” at the visual periphery—that is,
rather poor at telling colors apart if they fall on a retina position far
enough from the fovea so that the position contains mostly rods and very few
cones.
Related Topics