Chapter: Genetics and Molecular Biology: Protein Synthesis
Regulation of Ribosome Synthesis - Protein Synthesis
Regulation of Ribosome Synthesis
We already know that αr, during balanced exponential growth, is proportional to the growth rate. Let us consider the response of αrduring a transition between growth rates. Originally this type of data was sought in an effort to place constraints on the possible feedback loops in the ribosome regulation system. By analogy to electronic circuits, the response of the ribosome regulation system could be highly informative. The most straightforward growth rate shift is generated by addition of nutrients supporting faster growth. The resulting response in αr is a quick increase to the αr characteristic of the new growth conditions (Fig. 7.26). Superimposed on this shift are small oscillations. The existence of these oscillations shows that a number of cellular components are involved in the regulatory system. The ten-minute period of the oscilla-tions suggests that at least one of the regulating components is not a
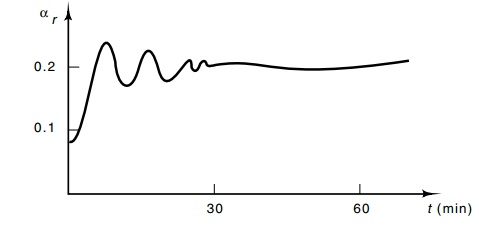
Figure 7.26 The response ofαr, to a growth rate increase.
In addition to regulating ribosome synthesis in response to differing growth rates, bacterial cells, and likely others as well, display a second type of ribosome regulation. This is the stringent response, in which the synthesis of ribosomal RNA and tRNA completely stops if protein synthesis stops. One obvious question about ribosome regulation is whether the stringent response itself is joined to the growth rate re-sponse system. Studies have shown that mutants in the stringent system, which are called relaxed, regulate their ribosome levels the same as wild-type cells as growth rate is varied. Consequently, these two systems are separate, as is verified by more recent experiments by Gourse.
The halt in the accumulation of rRNA in amino acid-starved, strin-gent cells appears to result from decreased initiations by RNA polym-erase rather than from degradation of rRNA or blockage of elongation by RNA polymerase. This is not surprising in light of what we have already discussed about regulation of DNA synthesis. One demonstra - tion of this fact is that no radioactive rRNA is synthesized if amino acids,
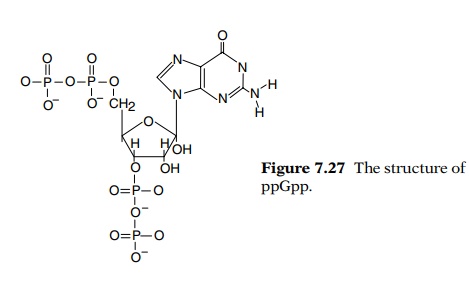
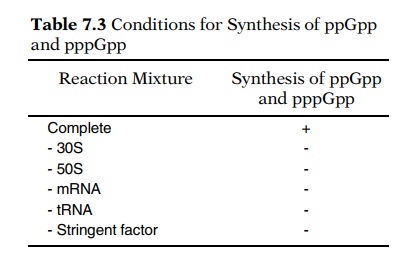
During the stringent response, guanosine tetra- and pentaphosphate, ppGpp (Fig. 7.27) and pppGpp, accumulate in amino acid-starved, stringent cells but not in amino acid-starved, relaxed cells. These com-pounds may directly interfere with transcription of the ribosomal RNA genes. In vitro experiments show that synthesis of ppGpp requires a particular protein, ribosomes, messenger, and an uncharged tRNA corresponding to the codon of messenger in the A site of a ribosome (Table 7.3). The required protein is normally rather tightly associated with the ribosomes and is the product of a gene called relA.
Related Topics