Chapter: Psychology: The Brain and the Nervous System
The Cerebral Cortex: Projection Areas
Projection
Areas
Many researchers divide the
cortex into three broad types of tissue. Sensory
areas receive and interpret information from the eyes, ears, and other
sense organs. The motor areas control
our behaviors—our movements through the world. The remaining areas are
typically called association areas
and are said to be involved in the complex processes broadly referred to as
thinking.
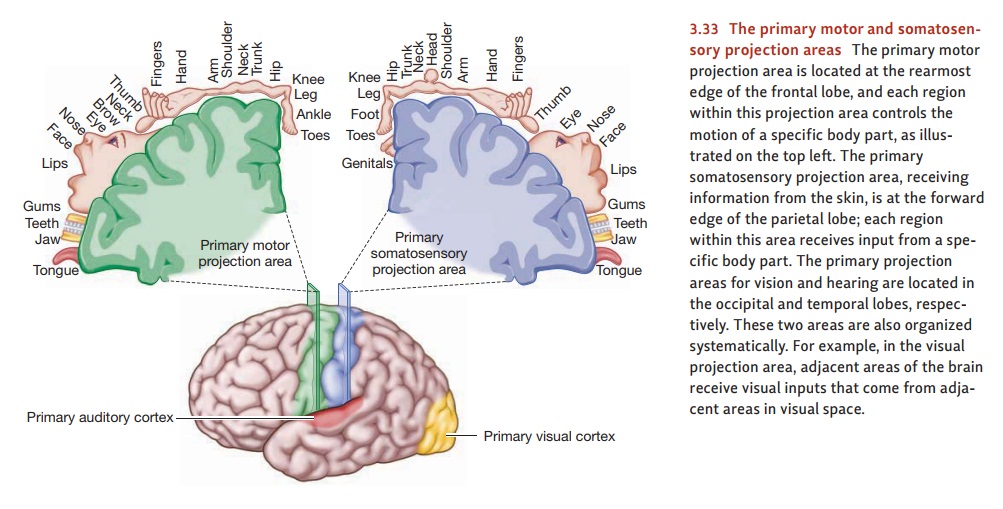
The sensory areas are divided by
modality, and so the brain region needed for vision is located in the occipital
cortex, and the region needed for hearing is in the temporal cortex (Figure
3.33). Within each of these modality-specific areas, we find projectionareas—areas where the brain
tissue seems to form a “map” of the sensory information.(The term projection is borrowed from mapmaking.)
Thus, for the visual projection
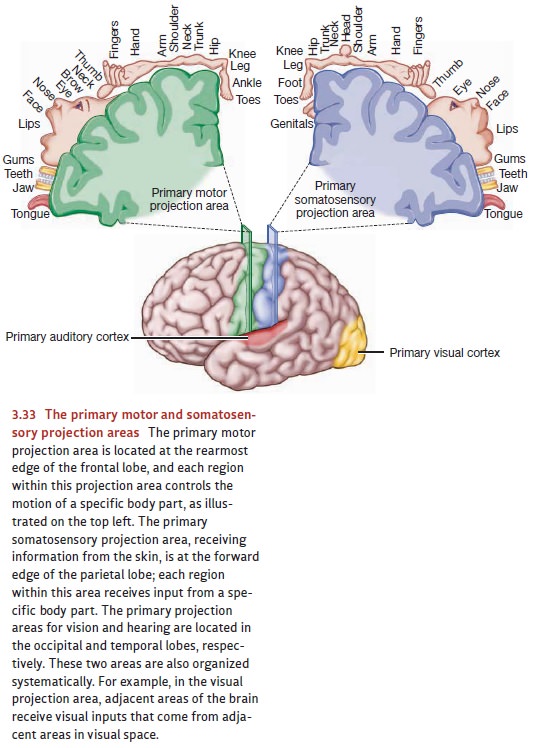
areas, adjacent sites in the
brain represent adjacent locations in the external world. For the auditory
projection areas, adjacent sites in the brain represent similar pitches.
As it turns out, the brain has
multiple projection areas—multiple maps—for each sense modality. The term primary projection area is therefore
used to designate the ini-tial receiving station for information arriving from
the sense organs—and so there’s a primary projection area for vision, one for
hearing, and one for information arriving from the skin.
We also find projection areas in
the motor areas of the cortex. In that case, adjacent sites in the brain
usually represent adjacent parts of the body. And there, too, we find a primary
projection area—the departure point
for signals that exit the cortex and ultimately result in muscle movement.
PRIMARY MOTOR AREAS
The discovery of the primary
motor projection area dates back to the 1860s, when investigators began to
apply mild electric currents to various portions of the cortex of anesthetized
animals. The effects were often quite specific. Within the frontal lobe, stimulating
one point led to a movement of the forelimb; stimulating another point made the
ears prick up. These early studies also provided evidence for the pattern of contralateral control, in which
stimulating the left hemisphere led to movements onthe right side of the body
and stimulating the right hemisphere caused movements on the left.
Similar results have been
obtained with humans. Canadian neurosurgeon Wilder Penfield, for example,
collected data from his patients who were undergoing surgery for epilepsy. The
surgery was intended to remove diseased tissue; and, as is common in
neurosurgery, the patients were awake during the procedure. (Because the brain
itself has no pain receptors, neurosurgery is often performed with only local
anesthesia that is required just to allow the surgeon to penetrate the scalp
and skull.) In the surgeries, Penfield confirmed that stimulating the motor
area in the frontal lobe led to movement of specific body parts—much to the
surprise of the patients, who had no sense of willing the action or performing
it themselves.
Systematic exploration persuaded
Penfield that for each portion of the motor cortex, there was a corresponding
part of the body that moved when its cortical counterpart was stimulated. These
findings are often summarized with a “motor homunculus,” a schematic picture
showing each body part next to the bit of the motor projection area that
controls its movement (Figure 3.33, top left).
As Figure 3.33 makes plain,
equal-sized areas of the body are not controlled by equal amounts of cortical
space. Instead, the parts that we can move with the greatest precision (e.g.,
the fingers, the tongue) receive more cortical area than those over which we
have less control (e.g., the shoulder, the abdomen). Evidently, what matters is
function—the extent and complexity of that body part’s use (Penfield &
Rasmussen, 1950).
PRIMARY SENSORY AREAS
Methods similar to Penfield’s
revealed the existence of sensory projection areas. The primary somatosensory projection area is directly behind the
primary motor pro-jection area, in the parietal lobe (see Figure 3.33, top
right). This area is the initial receiving area for sensory information
arriving from the skin senses. Patients stim-ulated at a particular point in
this area usually report tingling somewhere on the opposite side of their
bodies. (Less frequently, they report experiences of cold or warmth.) The
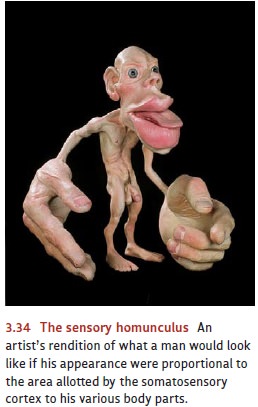
somatosensory projection area resembles its motor counterpart in several ways.
First, it shows an orderly projection pattern in which each part of the body ’s
surface sends its information to a particular part of the cortical
somatosen-sory area. Second, the assignment of cortical space is not in
proportion to the size of each body part. Instead, the space corresponds to the
sensitivity of each region; the parts of the body that are most sensitive to
touch (such as the index finger, lips, and tongue) receive more cortical space
(Figure 3.34). Finally, sensation—like motor control—is contralateral. That is,
sensory information from each part of the body proceeds to the brain hemisphere
on the side opposite to it—so that (for example) sensations from the right
thumb arrive in the left hemisphere, sensations from the left shoulder are sent
to the right hemisphere, and so on. (Information from the trunk of the body
close to the body ’s midline is represented in both hemispheres.)
The brain has similar primary
projection areas for vision and for hearing, and they’re located in the
occipital and temporal lobes, respectively (see Figure 3.33). Patients
stimulated in the visual projection area report optical experiences, vivid
enough but with little form or meaning—flickering lights, streaks of color.
When stimulated in the auditory area, patients hear things—clicks, buzzes,
booms, and hums.
As we noted earlier, the visual
and auditory areas are—like the somatosensory area—well organized spatially. In
the occipital lobe, especially the area known as the visual cortex, adjacent
brain areas represent adjacent locations in visual space. In the temporal
lobes, adjacent areas represent similar ranges of pitch. The visual area also
respects the principle of contralateral input: Objects seen (by either eye) to
the left of a person’s overall line of sight are processed by the right visual
area, and objects seen on the right are processed by the left visual area. The
auditory projection area, in contrast, provides a rare exception to the brain’s
contralateral wiring, because both cerebral hemispheres receive input from both
ears.
Related Topics