Chapter: Psychology: The Brain and the Nervous System
Propagation of the Action Potential
Propagation of
the Action Potential
So far, we have considered the
ion flow at just one point on the neuron’s membrane: When the membrane is
perturbed at that location, the ion channels there open, sodium ions rush in,
and the cell interior at that region briefly loses its negative charge. We said
earlier, though, that the neuron doesn’t just respond to an input; it also transmits this response from one end of
the neuron to other. Specifically, the neuron sends a signal down its axon,
where it will eventually launch a new series of events triggering the next neuron. How does this transmission
work?
As we’ve just noted, when a
neuron’s membrane is disturbed, it is briefly depolarized— that is, it loses the electrical charge that normally
exists across the membrane. This depolar-ization takes place at a particular
location; but it spreads because depolarization at one point on the membrane
causes other nearby ion channels to open, and so sodium rushes into the cell at
those locations as well. Of course, the resting potential is quickly restored
at those locations, but—for a brief
moment—a new portion of the cell’s membrane has beendepolarized. This
depolarization causes the next set of ion channels to open, which causes the
next set to open, and so on in a domino-like sequence as the action potential
at one site triggers an action potential at the next. In this way, the
depolarization moves down the entire length of the axon and throughout the rest
of the neuron as well. This sequence of events is known as the propagation of the action potential.
The whole thing is like a spark traveling along a fuse—except that whereas the
fuse is consumed by the spark, the ion channels rapidly reclose and the
membrane restores itself (i.e., reestablishes the resting potential; Figure
3.11).
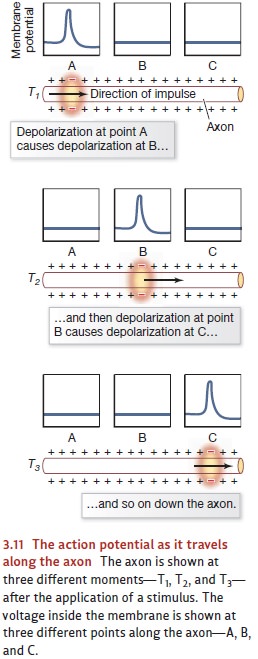
One might worry that this process
could continue infinitely—as one region of the membrane depolarizes its
neighbor, which in turn causes a new depolarization of the first region, which
then depolarizes the neighbor again, and on and on. This back-and-forth
disruption is prevented by the refractory period at each area of the mem-brane.
Thanks to the refractory period, the area of membrane that was depolarized
first is unresponsive when, a moment later, its neighbor is depolarized. As a
result, the action potential is propagated in one direction only and thus works
its way down the axon.
The flow of ions in or out of the
neuron is, as we’ve said, quite rapid. Even so, the propagation of the action
potential is surprisingly slow—it travels at about 1 meter per second, roughly
the walking speed of an average adult. If this was top speed for neural
signals, it would be disastrous for most organisms; fast-paced actions would be
impossi-ble. This problem is solved, however, by an anatomical feature we’ve
already mentioned: the myelin layers that wrap around an axon and—crucially—the
gaps between the myelin wrappers.
If an axon is myelinated, ions
can move into or out of the axon only at the nodes of Ranvier. At all other
locations, the axon is enclosed within its myelin wrapper, and this blocks ion
flow. In essence, therefore, the action potential has to skip from node to
node; and thanks to these jumps, it moves relatively quickly: Myelinated axons
can propagate their action potentials at speeds up to 120 meters per second
(about 260 miles per hour).
To appreciate the huge importance
of the myelin, consider the deficits someone suf-fers when myelination breaks
down in the brain. This happens in multiple
sclerosis(MS), a disease in which the body’s immune system mistakenly regards
the myelin as an intruder and attacks it. The manifestations of MS are variable
but severe, and they can include such serious maladies as total blindness and paralysis.
Related Topics