Chapter: The Massage Connection ANATOMY AND PHYSIOLOGY : Nervous System
Control of Posture and Movement
Control of Posture and Movement
The contraction of skeletal muscles ultimately depends on the pattern and discharge rate of the lower motor neurons supplying the muscle. These motorneurons, the final common pathway, are bombarded by impulses from thousands of neurons that synapse with it in the spinal cord and brainstem. Many synapses are from neurons in the same seg-ment. Some neurons that synapse are from the seg-ments above and below. There are other neurons that descend from the brainstem and cerebral cor-tex. It is the integrated activity of all these inputs that regulate posture and make coordinated move-ments possible.
The activity of the input achieves three functions:
(1) they produce voluntary movement; (2) they initiate adjustments to the posture to provide stability when parts move; and (3) they coordinate various muscle groups to make movements precise and smooth.
The motor cortex of the brain plans the patterns of voluntary activity and conveys it to the final com-mon pathway via the corticospinal and corticobul-bar tracts. Thecerebellum coordinates and makesthe movements smooth as the activity planned by the motor cortex; visual input, input from the inner ear, as well as input from proprioceptors are conveyed to it. The basal ganglia help maintain muscle tone and participate in automatic movements. Constant feed-back from the muscle and joints help the basal gan-glia and part of the cerebellum adjust the commands sent by the cortex.
GENERAL PRINCIPLES OF THE CONTROLOF VOLUNTARY MOVEMENTS
The commands for voluntary movement originate in the cortical association areas (see Figure 5.44). The movements are planned in the cortex, as well as the basal ganglia and part of the cerebellum. The com-mands are then relayed via the corticospinal tracts (from the cortex to the spinal nerves) and the corti-cobulbar tracts (from the cortex to the cranial nerves) to the lower motor neuron that supplies the muscle. As the movement occurs, receptors in the muscle— the muscle spindle, Golgi tendon organ, joint recep-tors, and those in the skin—are stimulated. This feed-back information is relayed back via sensory nerves to the cerebellum and the motor cortex and the move-ments are adjusted to make it smooth and precise.
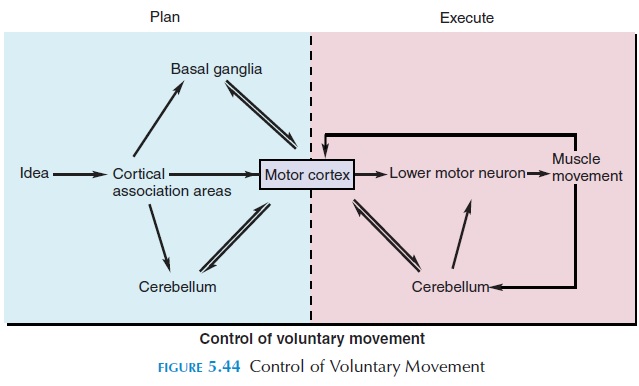
The neurons from the cerebellum project to the brainstem, from which they descend to the lower mo-tor neurons via the rubrospinal, reticulospinal, tec-tospinal, and vestibulospinal tracts.
THE MOTOR CORTEX
The motor cortex is located in the precentral gyrus. Similar to the sensory cortex, the various parts of the body are represented in the cortex, with the feet at the superior aspect of the gyrus and the face on the inferior and lateral aspect (see Figure 5.45). The fa-cial area is represented in both sides; however, the rest of the body is represented on one side, with the left cortex representing the right half of the body and the right cortex the left half. Again, similar to the sen-sory cortex, the size of cortical representation is in proportion to the number of motor units going to the muscle. This, in turn, correlates with the skill with which the part is used for fine, precise, voluntary movements. For example, although the hands are small, the cortical representation is large. The lips, pharynx, and tongue required for speech also have a large representation.
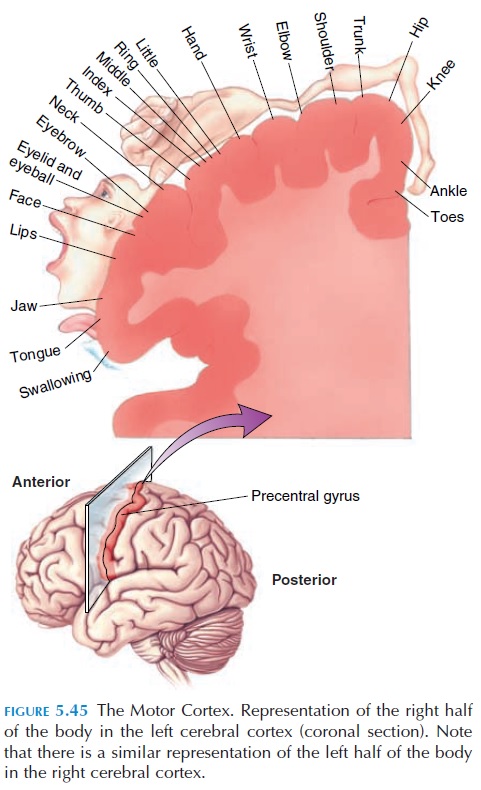
The corticospinal tracts and the corticobulbar tracts originate from the motor cortex. However, there are many other areas of the brain in the parietal lobe and elsewhere that participate in motor function.
Motor Pathway
The neurons responsible for voluntary control project from the motor cortex to the nerves supplying the muscles in question via direct, pyramidal, or corti-cospinal/corticobulbar pathways. They are calledpyramidal pathways because the axons form pyra-midlike bulges, the pyramids, in the medulla. Axons of neurons originating in the cortex descend to the medulla (see Figure 5.46). As they pass near the thal-amus, all the fibers lie close together in the region of the internal capsule.The sensory tracts are also in close proximity in this region.
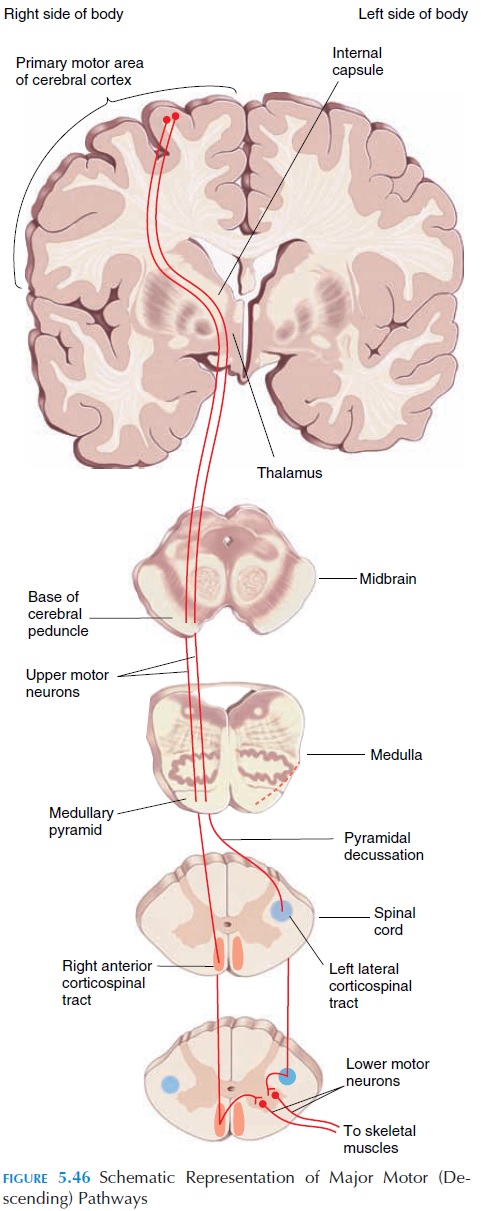
On reaching the medulla, most fibers of the corti-cospinal tract cross over to the opposite side, descend as the lateral corticospinal tract, and synapse with the respective motor neuron. The neurons in the cor-ticospinal tract are the upper motor neurons. Some axons descend on the same side as the anterior cor-ticospinal tract and cross over at the spinal segment where they synapse with lower motor neurons (the neurons that directly supply the muscles).
In this way, axons from one side control the muscles of the opposite side of the body. Just like the corticospinal tracts, the axons of upper motor neurons that control skeletal muscles in the head form the corticobulbartracts. They synapse with lower motor neurons thatexit via the cranial nerves.
In addition to the direct or pyramidal pathways, there are indirect or extrapyramidal motor path-ways. These tracts are complex and involve impulsesfrom the basal ganglia, limbic system, cerebellum, thalamus, and reticular formation, etc. Some tracts are the rubrospinal, tectospinal, reticulospinal, and vestibulospinal tracts.
LESIONS AND MUSCLE TONE
If there is a lesion in the lower motor neuron, the muscle it supplies atrophies (becomes smaller). There is loss of muscle tone, resulting in flaccidity. No re-flexes can be elicited because the muscle cannot be stimulated.
If there is a lesion in the upper motor neuron, the presentation is different because the lower motor neuron is intact. Also, the presentation will depend on which upper motor neuron is affected.
In a normal person, some descending tracts inhibit stretch reflexes and others stimulate; however, the in-hibitory effect is more prominent. If the corticospinal tract (has stimulatory effect) alone is injured, the muscle tone is diminished (hypotonic) and there is muscle weakness (paresis) rather than complete loss of movement.
If the extrapyramidal tracts are injured, the in-hibitory effect on the lower motor neuron is removed and the muscle tone is increased (hypertonic/spastic) and the reflexes are exaggerated. There is little muscle atrophy.
If the cerebellum or its projections are injured, there is incoordination of movement.
OTHER POSTURE-REGULATING SYSTEMS
As can be seen in Figure 5.46, the posture regulating mechanisms are multiple and controlled at many lev-els. At the spinal cord level, sensory stimuli produce simple reflex responses.
· At the medullary level, antigravity reflexes are regulated (i.e., changes in tone of different groups of muscles according to the effects of gravity).
· At the midbrain level, locomotor reflexes are present (i.e., walking movements can occur).
· At the hypothalamus limbic system level, changes in motor function in relation to emo-tions are produced.
· At the cerebral cortex level, initiation of move-ments and movements in relation to memory and conditioned reflexes occur.
Lesions in any of these levels result in retention of the reflexes and function mentioned below the level.
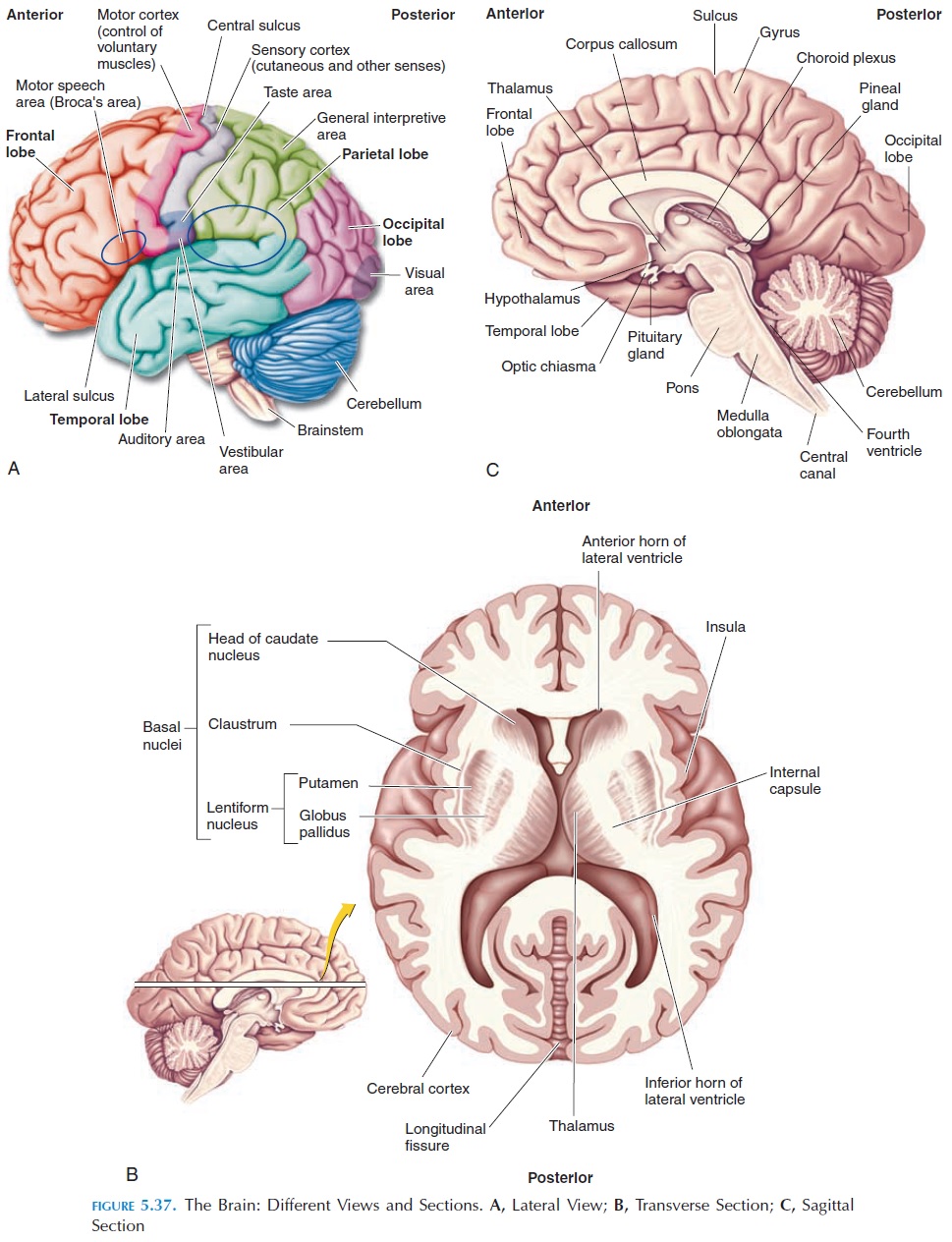
BASAL GANGLIA
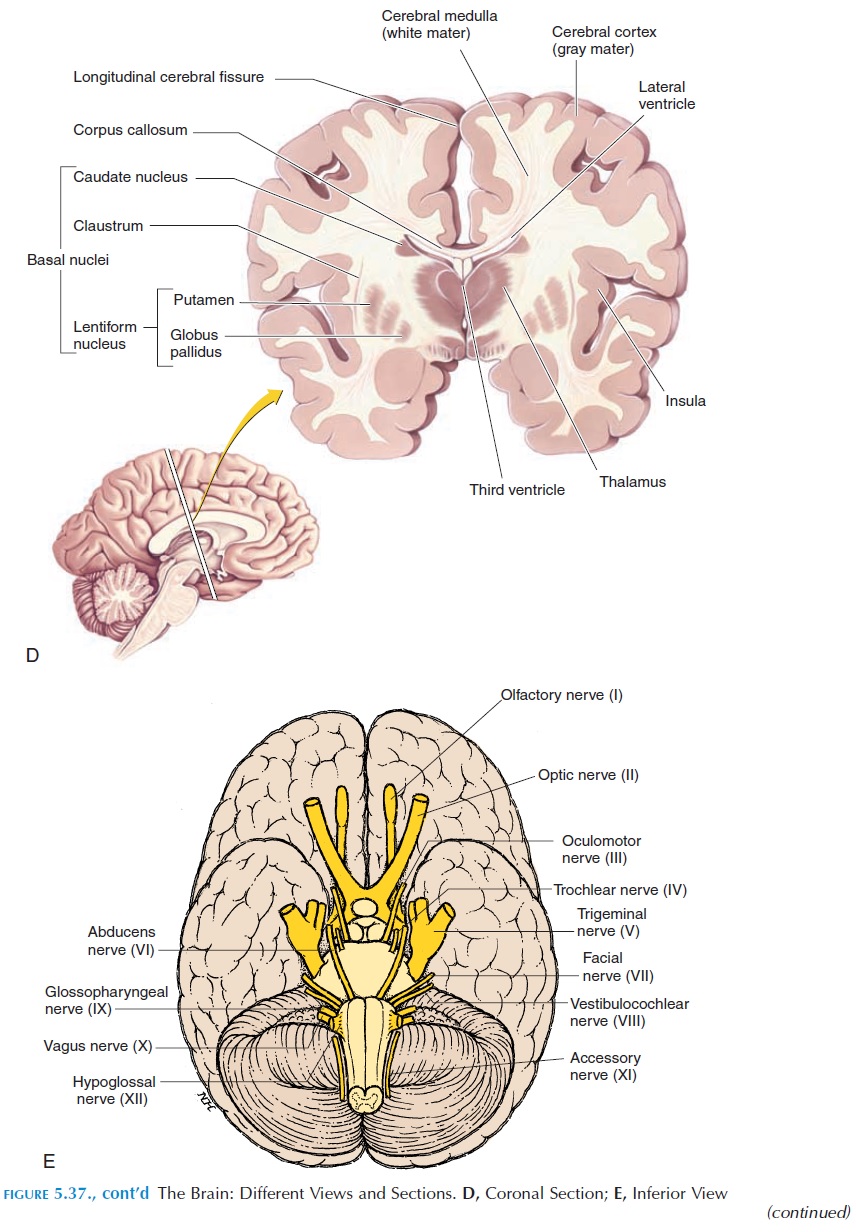
Deep to the cerebral cortex, there are many collec-tions of gray mater on both sides. The basal ganglia, or basal nuclei, are a group of these gray areas. The basal ganglia (Figure 5.37B) include the caudate nu-cleus, putamen, globus pallidus, the subthalamic nucleus, and the substantia nigra.
The basal ganglia have numerous connections. A major input is from various parts of the cerebral cor-tex and the thalamus. The different regions of the basal ganglia are extensively interconnected too. The basal ganglia, in turn, send efferents to the cortex via the thalamus and other areas.
The major function of the basal ganglia is its role in planning and programming movement. Its role can be better examined in animals and people with lesions in this region. Basal ganglia lesions are char-acterized by involuntary movement. Some move-ments and dysfunctions are described in Symptoms and Signs of Lesions in the Basal Nuclei.
CEREBELLUM
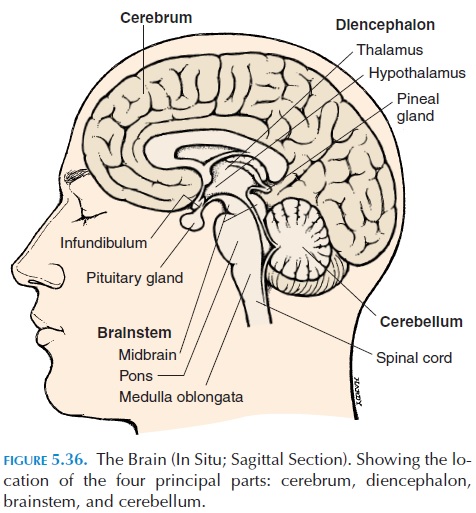
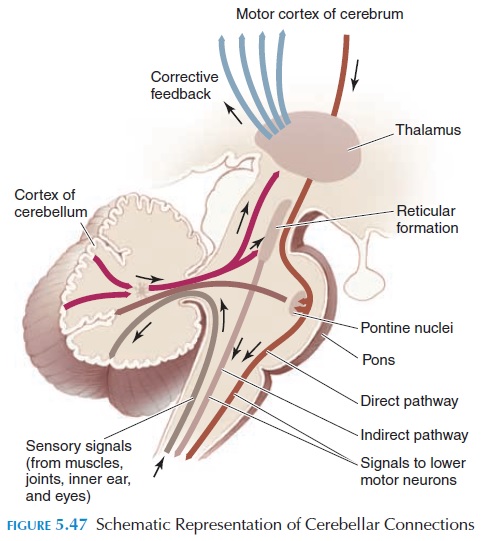
Another important region involved in posture and movement is the cerebellum (Figures 5.36 and 5.37). The cerebellum is required for learning and perform-ing rapid, coordinated, and skilled movements. It lies posterior to the brainstem; numerous tracts enter and leave in the brainstem and receive important input from proprioceptors located over the entire body. It also receives input from the vestibular apparatus (see below). The motor cortex and basal ganglia send im-pulses through the pontine nuclei that inform the cere-bellum of the motor plan. The cerebellum compares the motor plan with what is actually happening (feed-back from proprioceptors, vestibular apparatus, and eyes) and smoothes and coordinates the movement by sending impulses to the motor cortex (via the thala-mus) and the nuclei in the brainstem (see Figure 5.47).
Lesions of the cerebellum produce pronounced ab-normalities when an individual begins to move. The individual has ataxia—incoordination as a result of errors in the rate, range, force, and direction of movement. The individual has a drunken gait and in-voluntary movements and tremors when she or he in-tends to do something. Typically, every movement is performed in slow motion, as if every component of the movement has been dissected out and done one at a time. Cerebellar problems, however, do not affect the sensory system.
VESTIBULAR APPARATUS
This paired organ is part of the inner ear (see Figure 5.48) and sensations produced here are conveyed to the brain along with that of hearing via the vestibulo-cochlear nerve. Because it is directly related to equi-librium and balance, it is discussed in this section.
The vestibular apparatus, on each side, consists of three circular canals. Each canal lies perpendicular to each other and is enclosed in a bony labyrinth. To visualize this, place a half-open book, vertically on a table. Then each canal would lie along the two halves of the book, with one canal flat on the table. The canals are interconnected and filled with fluid. At the point where they meet, the canal is expanded to form the ampulla, which, in turn, is connected to fluid-filled structures called utricle and saccule.The am-pulla of the canals, the utricle and saccule have re-ceptors that respond to movement.
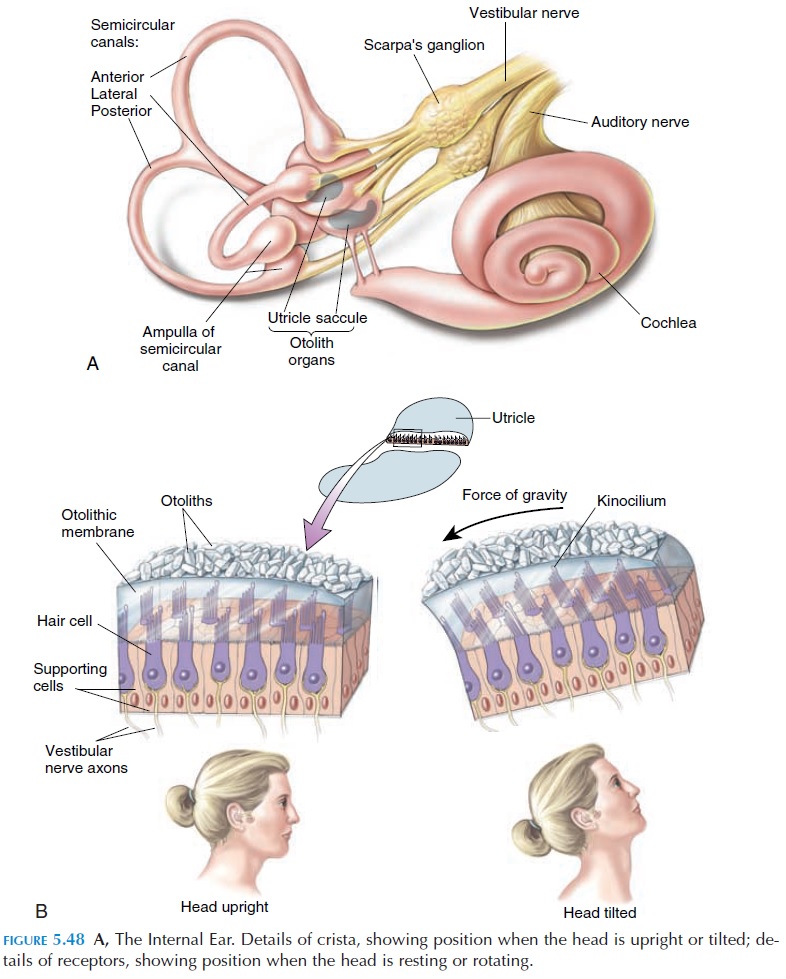
The receptors in the utricle and saccule are found in a small, thickened area called the macula. The receptors are in the form of hair cells with cilia, surrounded by a glycoprotein membrane, the otolithic mem-brane. Calcium carbonate crystals, known as otoliths, are found on top of the membrane. When the head is tilted, the otoliths move as a result of gravity and pull on the hair cells, resulting in changes in membrane potential and impulse formation. The receptors in the utricle and saccule respond when the head moves for-ward or backward—linear acceleration.
The receptors in the canals respond maximally when the head is rotated—rotational acceleration. These receptors are located in small, elevated regions in the ampulla known as crista. Here, too, the recep-tors are in the form of hair cells, but they are covered by a gelatinous mass known as the cupula. Every time the head is moved, the fluid in the semicircular canals is set in motion. Depending on which side of the head is turned, fluid in specific semicircular canals of the two sides move, causing the cupula to move in turn and generate impulses in the receptors.
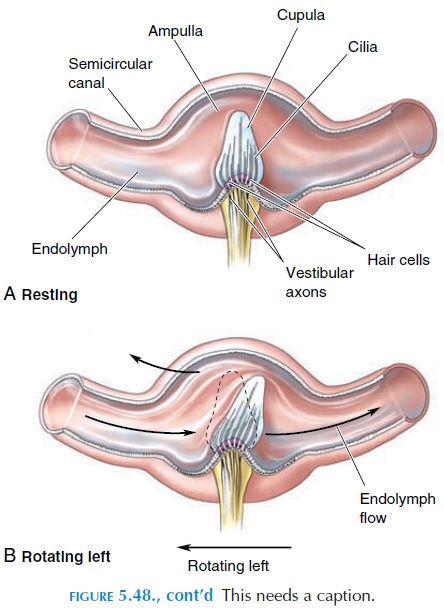
The impulses travel to the brainstem where some descend as the vestibulospinal tract that affects lower motor neurons. Certain impulses enter the cerebel-lum to give information about head movement. Cer-tain neurons take information to the cranial nerves that supply the eye to help the eye adjust to the move-ment; others take information to the motor cortex. In this way, the body is able to increase and decrease tone and maintain balance and equilibrium.
Because fluid has inertia, it continues to move even after stopping a rotational movement. This is the reason why individuals continue to feel giddy when they stop after turning rapidly. If the eyes were watched closely, rapid movements would be observed after stopping the movement (i.e., the eye movement persists for as long as the fluid continues to move in the canal).
The vestibular apparatus is important for orienta-tion in space. Orientation of the body is aided by vi-sual input, input from proprioceptors, and from touch and pressure receptors.
MOTOR SYSTEM LESIONS
Cortex and Corticospinal (Pyramidal) Lesions
Lesions of the cortex or corticospinal tract (e.g., stroke) result in muscle weakness without atrophy. Atrophy may ensue later because of disuse. The weakness is more in the extensors than the flexors in the upper limb, and so the upper limbs tend to be flexed. In the lower limb, it is the opposite, with the extensors being stronger. Stretch reflexes like the knee jerk tend to be brisk. There is dorsiflexion of the foot if the plantar response is elicited (positive Babinski’s sign). Because the tracts cross over at themedulla, the opposite half of the body is affected— hemiplegia. If the lesion is in the brainstem area,the functioning of cranial nerves that arise from there is also affected.
Spinal Cord Lesions
Spinal Shock
As soon as the spinal cord is injured or cut, it is fol-lowed by a period of spinal shock when all spinal reflex responses are depressed. This lasts for about two weeks in humans. The cause of spinal shock is uncertain.
With time, the spinal reflexes below the cut be-come exaggerated and hyperactive. It could be a re-sult of many reasons. One reason is the removal of the inhibitory effects of the higher motor centers. Also, the neurons become hypersensitive to the exci-tatory neurotransmitters. In addition, the spinal neu-rons may sprout collaterals that synapse with excita-tory input. Whatever the reason, the stretch reflexes are exaggerated and muscle tone increases. The first reflex response that comes back is a slight contrac-tion of the leg flexors and adductors in response to some painful stimuli.
The extent of disability depends on the level of the spinal cord that has been injured. It must be remem-bered that although the spinal cord has all the seg-ments—8 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 1 coccygeal—the length of spinal cord is shorter than the vertebral column and ends at level L1 and L2. Hence, injury below the second lumbar vertebra may affect only the muscles and dermatomes inner-vated by the sacral and coccygeal nerves.
If spinal cord injury occurs above the third cervi-cal spinal segment, other than the loss of voluntary movements of all the limbs, respiratory movements are affected as the phrenic nerve arising from C3, 4, 5 supplies the diaphragm. Loss of movement of all four limbs is known as quadriplegia. If the lesion is lower, only the lower limbs are affected, and this is termed paraplegia. If the nerves to only one limb are affected, it is referred to as monoplegia.
Other Complications of Spinal Cord Injuries
One common complication among people with spinal cord injuries is decubitus ulcer. Because vol-untary weight shifting does not occur, the weight of the body compresses the circulation to the skin over bony prominences, producing ulcers. These ulcers heal slowly and are prone to infection.
As a result of disuse, calcium from bones are reab-sorbed and excreted in the urine. This increases the incidence of calcium stones forming in the urinary tract. Paralysis of the muscles of the urinary bladder, in addition to stone formation, result in stagnation of urine and urinary tract infection.
When the spinal reflexes return, they are exagger-ated. For example, in a person with quadriplegia, the slightest of stimuli can trigger the withdrawal reflex and the stimulated limb flexes with flexion/extension responses from the other three limbs. With time, as a result of prolonged and repeated flexion, scar tissue may form in the limb and the limb becomes fixed in one position, known as contractures.
The function of the autonomic system below the level of lesion is also affected. Voluntary control is lost if the lesion is above the sacral segments, and reflex contractions of a bladder and rectum occur as soon as they get full. Bouts of sweating and blanching of the skin as a result of vasoconstriction of blood vessels may occur. Wide swings in blood pressure can occur as a result of imprecise blood pressure regulation.
Even though sexual reflexes are complex, with inte-gration at various levels, manipulation of the genitals in males can produce erection and even ejaculation.
Mass Reflex
Below the level of the injury, afferent stimuli can travel from one level to the other and even a slight stimulus to the skin can trigger many reflexes, such as emptying the bladder and rectum, sweating, and blood pressure changes. This is known as the massreflex. People with chronic spinal injuries use this re-flex to give them some degree of control over urina-tion and defecation. They can be trained to initiate these reflexes by stroking or pinching the thigh trig-gering the mass reflex intentionally.
RETICULAR ACTIVATING SYSTEMAND AROUSAL MECHANISMS
The various sensory impulses reach the cerebral cor-tex for perception and localization, as already ex-plained. En route, these nerves send collaterals to the reticular activating system (RAS), a network of neu-rons located in the brainstem. This system is largely responsible for the conscious, alert state of the body.
The reticular formation, or RAS, is a network of small neurons located at the center of the medulla and midbrain. It is a complex network with varied functions and is the site where the cardiac, vasomo-tor, and respiratory centers are located. The reticular formation also communicates with sensory pathways and plays a role in motor reflexes. Only its function in arousal and sleep are elaborated here.
The reticular formation has input not only from the general sensory tracts, but also from taste, smell, visual, and hearing sensations. Fibers from the retic-ular formation go to every part of the cortex, some via the thalamus, and it is the activity of these fibers that is largely responsible for the electrical activity of the cortex. When one is asleep, the reticular activat-ing system is in a dormant state. But it becomes ac-tive when there is any form of sensory input and the electrical activity of the brain increases—the arousal reaction. Many theories have been presented to ex-plain sleep. It is believed that sleep may be a result of fatigue of the various synapses in the brain. Secretion of certain chemicals in the brain has also been thought to induce sleep.
Sleep consists of different levels with two compo-nents: nonrapid eye movement (NREM) sleep and rapid eye movement (REM) sleep. The different lev-els of sleep can be identified by the characteristic ap-pearance of waves in electroencephalograph (EEG) recordings.
LEARNING AND MEMORY
Learning is the ability to acquire new skills or knowl-edge, and memory is the ability to retain what is learned. Many areas of the brain, including the asso-ciation areas, parts of the limbic system, thalamus, and hypothalamus, are believed to be involved in these processes. Although there are no complete ex-planations for how we learn or how memory is stored, it has been shown that neurons have the abil-ity to change in response to stimuli from internal and external environments. This ability, referred to asplasticity, is associated with changes in productionof specific proteins by neurons and formation of new dendrites and new synapses and neuronal circuits. Interestingly, it has been shown that areas of the cere-bral cortex that are not used become thinner and those areas used extensively become larger.
MENINGES, CEREBROSPINALFLUID, AND ITS CIRCULATION
The spinal cord and brain are delicate structures that need protection. They are so delicate that the brain can be scooped out of the skull with a spoon. They are protected, to a large extent, by the body cover-ing—the vertebral column and the tough connective tissue sheaths—the meninges.
The dura mater in the cranial cavity, unlike that in the spinal region, is a double-layered structure with the outer layer adhering to the periosteum and the in-ner layer following the contours of the brain. In some regions of the cranial cavity, the two layers are sepa-rated to enclose the dural sinus (see Figure 5.49) containing venous blood. This blood eventually drains into the internal jugular vein. In some regions of the cranial cavity, the dura also forms thick septae that separate major structures on the surface of the brain. For example, a thick layer of dura, the falxcerebri, separates the right and left cerebral hemi-spheres and the tentorium cerebelli separates the cerebrum from the cerebellum.
The arachnoid and the pia mater are similar in structure to that around the spinal cord .
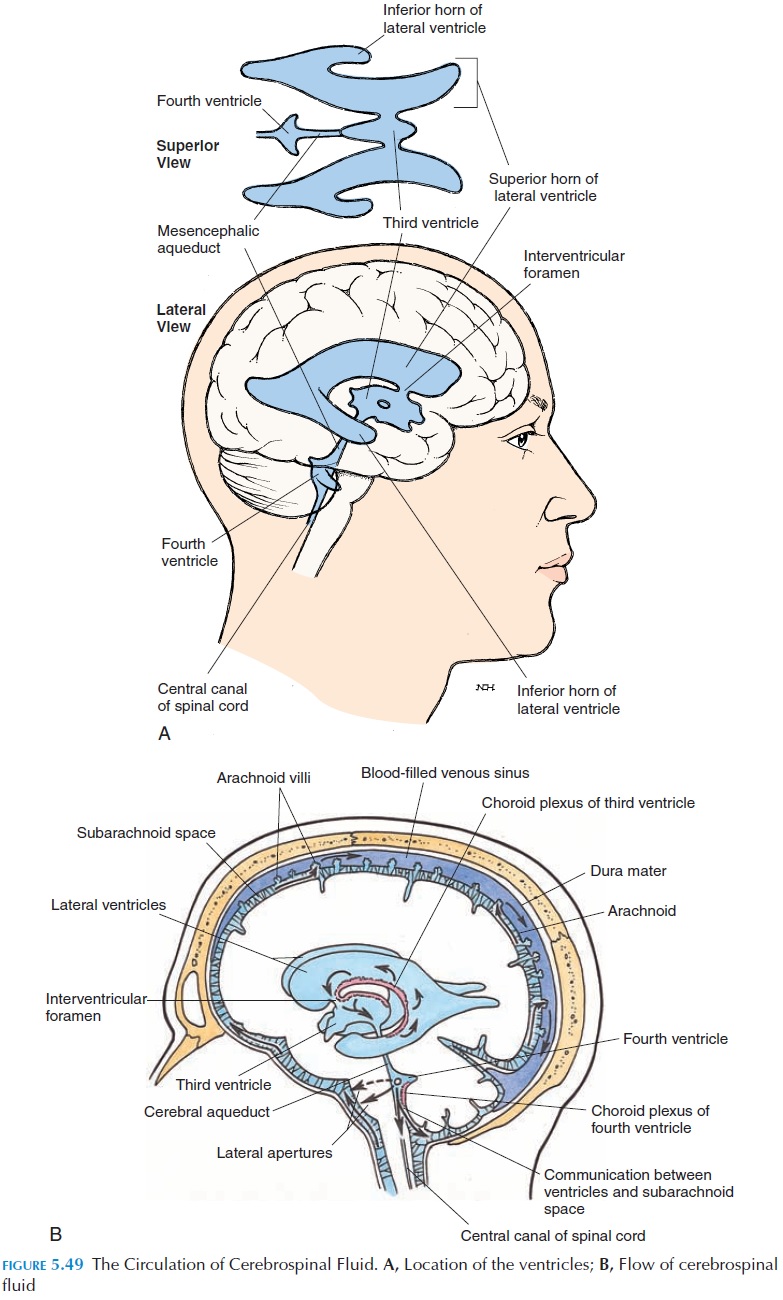
For further protection, the CNS floats in a cushion of fluid known as the cerebrospinal fluid (CSF). This fluid is manufactured from blood by specialized secretory structures, the choroid plexus, located in-side the brain. The CSF then flows through four widened chambers and narrower channels located inside the brain and exit into the subarachnoid space through openings located in the brainstem. From the subarachnoid space, the CSF flows into the large veins draining the brain (Figure 5.49B).
The four widened chambers inside the brain are known as ventricles (Figure 5.49A). The two lateralventricles are large and located in the cerebrum. Theyextend from the frontal lobe anteriorly to the occipital lobe. In the region of the parietal lobe, an inferior ex-tension of the cavity projects into the temporal lobe.
The lateral ventricles communicate with another cavity, the third ventricle, that is located in the mid-line between the thalamus and hypothalamus. The third ventricle narrows inferiorly and opens into an-other widened area, the fourth ventricle, that is lo-cated in the pons and medulla anterior to the cerebel-lum. Three openings located in the fourth ventricle allow the CSF to flow into the subarachnoid space.
The CSF is a clear, watery fluid; the volume is about 150 mL (5 oz). The composition is similar to that of plasma without the proteins and cells. The normal cir-culation of CSF and its volume and pressure is a bal-ance between the rate at which it is produced by the choroid plexus and absorbed into the veins.
BLOOD SUPPLY TO THE BRAIN
Related Topics