Chapter: Biochemistry: Photosynthesis
Chloroplasts Are the Site of Photosynthesis
Chloroplasts Are the Site of
Photosynthesis
It is
well known that photosynthetic organisms, such as green plants, convert carbon
dioxide (CO2) and water to carbohydrates such as glucose (written
here as C6H12O6) and molecular oxygen (O2).
6CO2
+ 6H2O - > C6H12O6 + 6O2
The
equation actually represents two processes. One process, the oxidation of water
to produce oxygen (the light reactions), requires light energy from the Sun.
The light reactions of photosynthesis in prokaryotes and eukaryotes depend on
solar energy, which is absorbed by chlorophyll
to supply the energy needed in the light reactions. The light reactions also
generate NADPH, which is the reducing agent needed in the dark reactions. The
other process, the Þxation of CO2 to give sugars (the dark
reactions), does not use solar energy directly but rather uses it indirectly in
the form of the ATP and NADPH produced in the course of the light reactions.
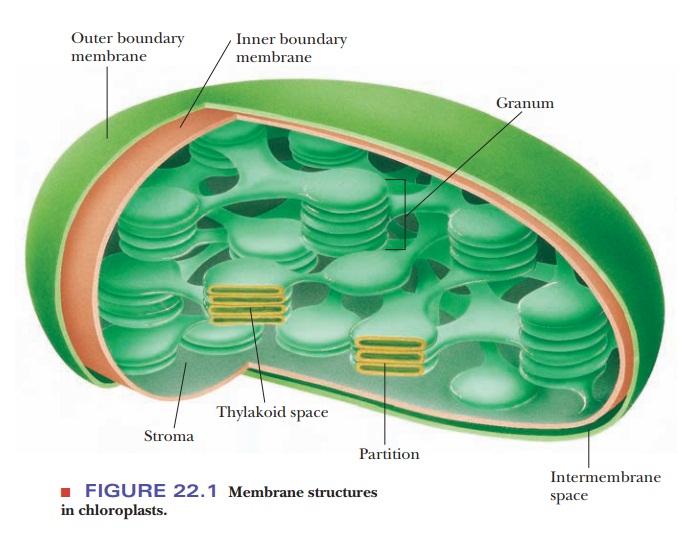
In
prokaryotes such as cyanobacteria, photosynthesis takes place in granules bound
to the plasma membrane. The site of photosynthesis in eukaryotes such as green
plants and green algae is the chloroplast
(Figure 22.1). Like the mitochondrion, the chloroplast has inner and outer
membranes and an intermembrane space. In addition, within the chloroplast are
bodies called grana, which consist
of stacks of flattened membranes called thylakoid
disks. The grana are connected by membranes called intergranal lamellae. The thylakoid disks are formed by the folding
of a third membrane within the chloroplast. The folding of the thylakoid
membrane creates two spaces in the chloroplast in addition to the intermembrane
space. The stroma lies within the
inner membrane and outside the thylakoid membrane. In addition to the stroma,
there is a thylakoid space within
the thylakoid disks themselves. The trapping of light and the produc-tion of
oxygen take place in the thylakoid disks. The dark reactions (also called
light-independent reactions), in which CO2 is fixed to
carbohydrates, take place in the stroma (Figure 22.2).

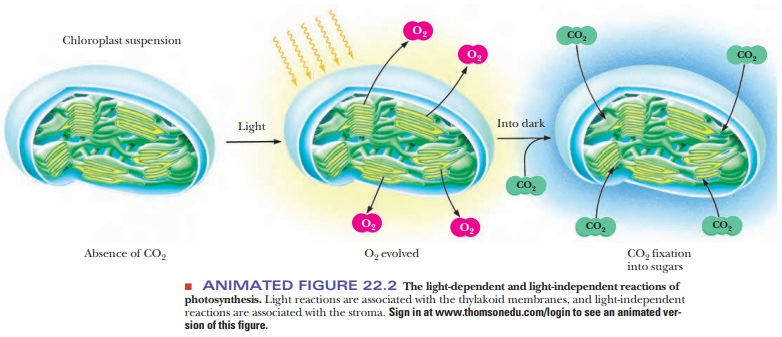
How does the structure of the chloroplast affect photosynthesis?
It is well established that the primary event in photosynthesis is the absorption of light by chlorophyll. The high energy states (excited states) of chlorophyll are useful in photosynthesis because the light energy can be passed along and converted to chemical energy in the light reaction. There are two principal types of chlorophyll, chlorophyll a and chlorophyll b. Eukaryotes such as green plants and green algae contain both chlorophyll a and chlorophyll b.
Prokaryotes such as cyanobacteria (formerly called blue-green
algae) con-tain only chlorophyll a.
Photosynthetic bacteria other than cyanobacteria have bacteriochlorophylls,
with bacteriochlorophyll a being the
most common. Organisms such as green and purple sulfur bacteria, which contain
bacterio-chlorophylls, do not use water as the ultimate source of electrons for
the redox reactions of photosynthesis, nor do they produce oxygen. Instead,
they use other electron sources such as H2S, which
produces elemental sulfur instead of oxygen. Organisms that contain
bacteriochlorophyll are anaerobic and have only one photosystem, whereas green
plants have two different photosystems, as we shall see.
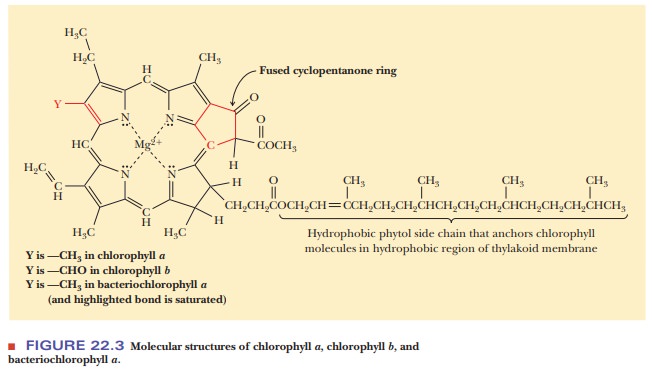
The structure of chlorophyll is similar to that of the heme group
of myoglo-bin, hemoglobin, and the cytochromes in that it is based on the
tetrapyrrole ring of porphyrins (Figure 22.3). The metal ion bound to the
tetrapyrrole ring is magnesium, Mg(II), rather than the iron that occurs in
heme. Another difference between chlorophyll and heme is the presence of a
cyclopentanone ring fused to the tetrapyrrole ring. A long hydrophobic side
chain, the phytol group, contains four isoprenoid units (five-carbon units that
are basic building blocks in many lipids;) and binds to the thylakoid membrane
by hydrophobic interactions. The phytol group is covalently bound to the rest
of the chlorophyll molecule by an ester linkage between the alcohol group of
the phytol and a propionic acid side chain on the porphyrin ring. The
difference between chlorophyll a and
chlorophyll b lies in the
substitution of an aldehyde group for a methyl group on the porphyrin ring. The
difference between bacteriochlorophyll a
and chlorophyll a is that a double
bond in the porphyrin ring of chlorophyll a
is saturated in bacteriochlorophyll a.
The lack of a conjugated system (alternating double and single bonds) in the
porphyrin ring of bacteriochlorophylls causes a significant difference in the
absorption of light by bacteriochlorophyll a
compared with chlorophyll a and b.
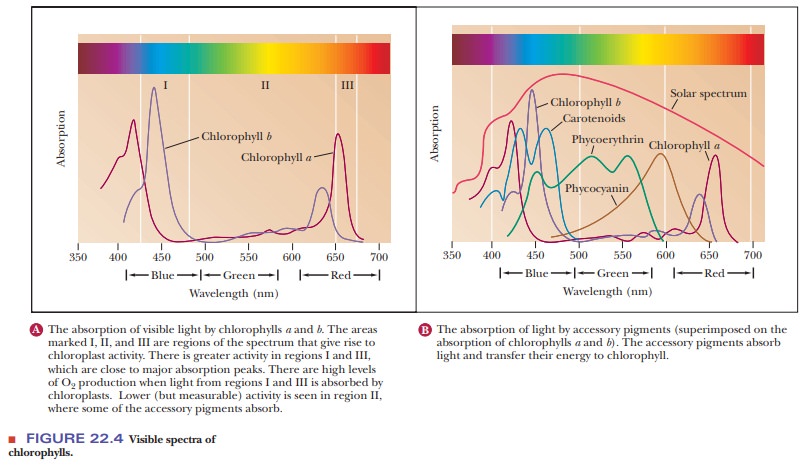
The absorption spectra of chlorophyll a and chlorophyll b differ slightly (Figure 22.4). Both absorb light in the red and blue portions of the visible spec-trum (600 to 700 nm and 400 to 500 nm, respectively), and the presence of both types of chlorophyll guarantees that more wavelengths of the visible spectrum are absorbed than would be the case with either one individually. Recall that chlorophyll a is found in all photosynthetic organisms that produce oxygen. Chlorophyll b is found in eukaryotes such as green plants and green algae, but it occurs in smaller amounts than chlorophyll a. The presence of chlorophyll b, however, increases the portion of the visible spectrum that is absorbed and thus enhances the efficiency of photosynthesis in green plants compared with cya-nobacteria. In addition to chlorophyll, various accessory pigments absorb light and transfer energy to the chlorophylls (Figure 22.4b). Bacteriochlorophylls, the molecular form characteristic of photosynthetic organisms that do not pro-duce oxygen, absorb light at longer wavelengths. The wavelength of maximum absorption of bacteriochlorophyll a is 780 nm; other bacteriochlorophylls have absorption maxima at still longer wavelengths, such as 870 nm or 1050 nm. Light of wavelength longer than 800 nm is part of the infrared, rather than the visible, region of the spectrum. The wavelength of light absorbed plays a critical role in the light reaction of photosynthesis because the energy of light is inversely related to wavelength.
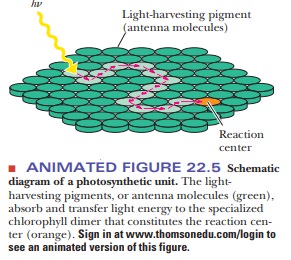
Most of the chlorophyll molecules in a chloroplast simply gather
light (antennae chlorophylls). All chlorophylls are bound to proteins, either
in antennae complexes or in one of two kinds of photosystems (membrane-bound protein complexes that carry out the
light reactions). The light-harvesting molecules then pass their excitation
energy along to a specialized pair of chlorophyll molecules at a reaction center characteristic of each
photosystem (Figure 22.5). When the light energy reaches the reaction center,
the chemical reactions of photosynthesis begin. The different environments of
the antennae chlorophylls and the reaction-center chlorophylls give different
properties to the two different kinds of molecules. In a typical chloroplast,
several hundred light-harvesting antennae chlorophylls are present for each
unique chlorophyll at a reaction center. The precise nature of reaction centers
in both prokaryotes and eukaryotes is the subject of active research.
Summary
In eukaryotes, photosynthesis takes place in chloroplasts. The
light reac-tions take place in the thylakoid membrane, a third membrane in
chloro-plasts in addition to the inner and outer membrane.
The dark reactions of photosynthesis take place in the stroma, the
space between the thylakoid membrane and the inner membrane of the chloroplast.
The absorption of light by chlorophyll supplies the energy required
for the reactions of photosynthesis. Several different kinds of chlorophyll are
known. All have a tetrapyrrole ring structure similar to that of the
porphy-rins of heme, but they also have differences that affect the wavelength
of light they absorb.
This property allows more wavelengths of sunlight to be absorbed
than would be the case with a single kind of chlorophyll.
Related Topics