Chapter: Biochemistry: Photosynthesis
Dark Reactions of Photosynthesis Fix CO2
Dark Reactions of Photosynthesis
Fix CO2
The
actual storage form of the carbohydrates produced from carbon dioxide by
photosynthesis is not glucose but disaccharides (e.g., sucrose in sugarcane and
sugar beets) and polysaccharides (starch and cellulose). However, it is
customary and convenient to write the carbohydrate product as glucose, and we
shall follow this time-honored practice.
Carbon
dioxide fixation takes place in the stroma. The equation for the overall
reaction, like all equations for photosynthetic processes, is deceptively
simple.
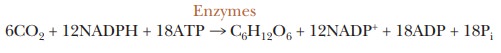
6CO2
+ 12NADPH + 18ATP - - (Enzymes)- > C6H12O6
+ 12NADP+ + 18ADP + 18Pi
The
actual reaction pathway has some features in common with glycolysis and some in
common with the pentose phosphate pathway.
The net
reaction of six molecules of carbon dioxide to produce one mol-ecule of glucose
requires the carboxylation of 6 molecules of a five-carbon key intermediate,
ribulose-1,5-bisphosphate, to form 6
molecules of an unstable six-carbon intermediate, which then splits to give 12
molecules of 3-phospho-glycerate. Of these, 2 molecules of 3-phosphoglycerate
react in turn, ultimately producing glucose. The remaining 10 molecules of
3-phosphoglycerate are used to regenerate the 6 molecules of ribulose-1,5-bisphosphate. The overall reaction
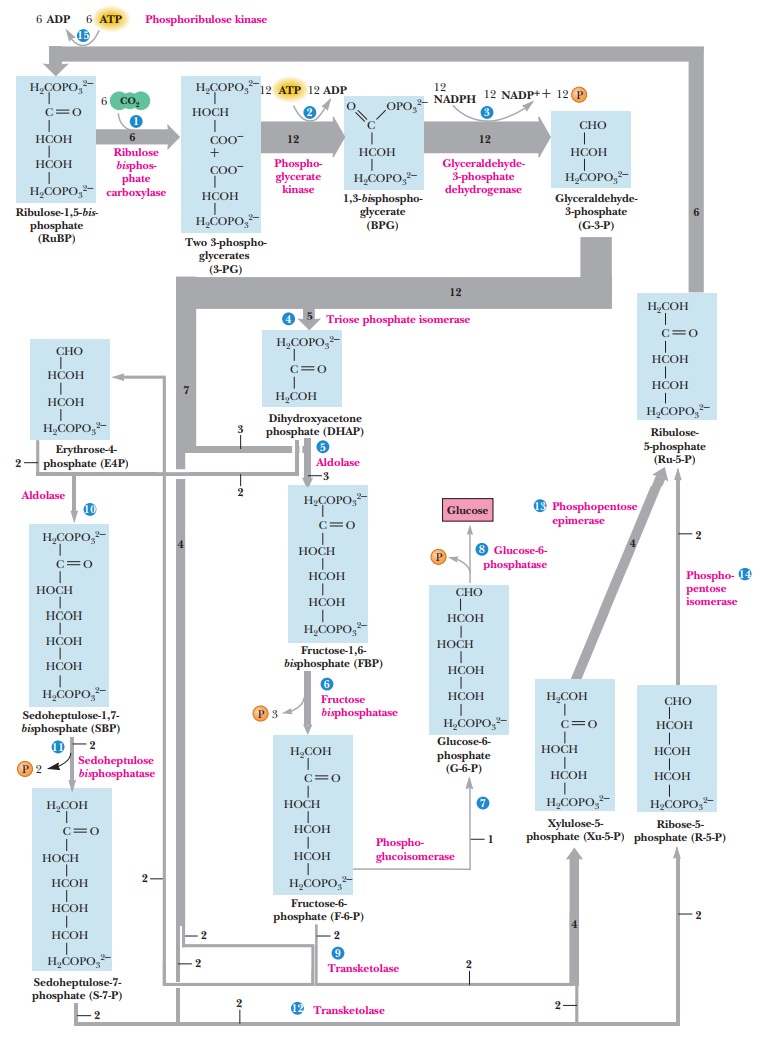
pathway is cyclic and is called the Calvin
cycle (Figure 22.15) after the scientist who first investigated it, Melvin
Calvin, winner of the 1961 Nobel Prize in chemistry.

What is the Calvin cycle?
The Þrst
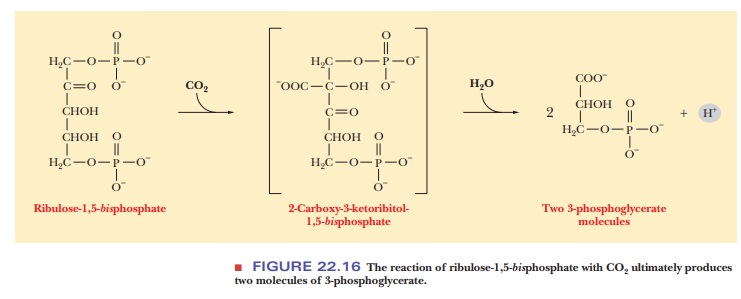
reaction of the Calvin cycle is the condensation of ribulose-1,5-bisphosphate with carbon dioxide to form
a six-carbon intermediate, 2-carboxy-3-ketoribitol-1,5-bisphosphate, which quickly hydrolyzes to give two molecules of
3-phosphoglycerate (Figure 22.16). The reaction is catalyzed by the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco).This enzyme is locatedon the stromal side of the
thylakoid membrane and is probably one of the most abundant proteins in nature,
as it accounts for about 15% of the total protein in chloroplasts. The
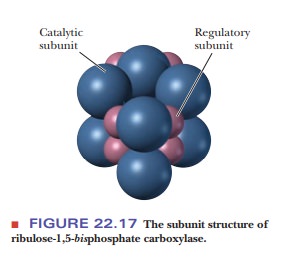
molecular weight of ribulose-1,5-bisphosphate
carboxylase/ oxygenase is about 560,000, and it consists of eight large
subunits (molecular weight 55,000) and eight small subunits (molecular weight
15,000) (Figure 22.17). The sequence of the large subunit is encoded by a
chloroplast gene,
The endosymbiotic theory for
the development of eukaryotes is consistent with the idea of independent
genetic material in organelles. The large subunit (chloroplast gene) is
catalytic, whereas the small subunit (nuclear gene) plays a regulatory role, an
observation that is consistent with an endosymbiotic origin for organelles such
as chloroplasts.
The
incorporation of CO2 into 3-phosphoglycerate represents the actual
fixation process; the remaining reactions are those of carbohydrates. The next
two reactions lead to the reduction of 3-phosphoglycerate to form
glyc-eraldehyde-3-phosphate. The reduction takes place in the same fashion as
in gluconeogenesis, except for one unique feature (Figure 22.15): the reactions
in chloroplasts require NADPH rather than NADH for the reduction of 1,3-bisphosphoglycerate to
glyceraldehyde-3-phosphate. When glyceraldehyde-3-phosphate is formed, it can
have two alternative fates: one is the produc-tion of six-carbon sugars, and
the other is the regeneration of ribulose-1,5-bisphosphate. Table 22.1 summarizes the reactions that take place
and indi-cates their stoichiometry.

The
formation of glucose from glyceraldehyde-3-phosphate takes place in the same
manner as in gluconeogenesis (Figure 22.15 and reactions 4 through 8 in Table
22.1). The conversion of glyceraldehyde-3-phosphate to dihydroxyacetone
phosphate takes place easily. Dihydroxyacetone phosphate in turn reacts with
glyceraldehyde-3-phosphate, in a series of reactions we have already seen, to
give rise to fructose-6-phosphate and ultimately to glucose. Because we have
already seen these reactions, we shall not discuss them again.
How is starting material regenerated in the Calvin cycle?
This process is readily divided into four steps: preparation, reshuffling, isomerization, and phosphorylation. The preparation begins with conversion of some of the glyceraldehyde-3-phosphate to dihydroxyacetone phosphate (catalyzed by triose phosphate isomerase). This reaction also functions in the production of six-carbon sugars. Portions of both the glyceraldehyde-3-phosphate and the dihydroxyacetone phosphate are then condensed to form fructose-1,6-bisphosphate (catalyzed by aldolase). Fructose-1,6-bisphosphate is hydrolyzedto fructose-6-phosphate (catalyzed by fructose-1,6-bisphosphatase). (See Figure 22.15. Reactions 4 through 6 in Table 22.1 are involved here.) With a supply of glyceraldehyde-3-phosphate, dihydroxyacetone phosphate, and fructose-6-phosphate now available, the reshufßing can begin.
Most of
the reactions of the reshuffling process are the same as ones we have already
seen as part of the pentose phosphate pathway. Consequently, we shall
concentrate just on the main outline of the process later because the results
are summarized in Figure 22.15 and Table 22.1. Reactions catalyzed in turn by transketolase, aldolase, and sedoheptulose bisphosphatase (Reactions 9 through 12 in Table 22.1) are the
reactions of rearrangement of carbon skeletons in the reshuffling phase of the
Calvin cycle.
The
isomerization step (reactions 13 and 14 in Table 22.1) involves the conversion
of both ribose-5-phosphate and xylulose-5-phosphate to ribulose-5-phosphate. Ribose-5-phosphate isomerase catalyzes
the conversion of ribose-5-phosphate to ribulose-5-phosphate, and xylulose-5-phosphate epimerase catalyzes
the conversion of xylulose-5-phosphate to ribulose-5-phosphate (Figure 22.15).
The reverse of both these reactions takes place in the pentose phosphate
path-way, catalyzed by the same enzymes.
In the
final step (reaction 15 in Table 22.1), ribulose-1,5-bisphosphate is regenerated by the phosphorylation of
ribulose-5-phosphate. This reaction requires ATP and is catalyzed by the enzyme
phosphoribulose kinase. The
reac-tions leading to the regeneration of ribulose-1,5-bisphosphate summarized in Table 22.1 give a net equation obtained
by adding all the reactions.
Taking
these points into consideration, we arrive at the net equation for the path of carbon in photosynthesis.
6CO2 + 18ATP + 12NADPH + 12H+ + 12H2O - - > Glucose + 12NADP+ + 18ADP + 18Pi
The
efÞciency of energy use in photosynthesis can be calculated fairly easily. The ∆G° for the reduction of CO2to
glucose is +478 kJ (+114 kcal) for each mole ofCO2 (see Question
37), and the energy of light of 600-nm wavelength is 1593 kJ mol−1
(381 kcal mol−1). We shall not explain in detail here how this Þgure
for the energy of the light is obtained, but it comes ultimately from the
equation E = hv. Light of wavelength 680 nm or 700 nm has lower energy than
light at 600 nm. Thus, the efÞciency of photosynthesis is at least (478/1593) -
> 100, or 30%.
Summary
In the dark reactions of photosynthesis, the fixation of carbon
dioxide takes place when the key intermediate ribulose-1,5-bisphosphate reacts with carbon dioxide to produce two molecules of
3-phosphoglycerate. This reaction is catalyzed by the enzyme ribulose-1,5-bisphosphate carbox-ylase/oxygenase
(rubisco), one of the most abundant proteins in nature.
The remainder of the dark reaction is the regeneration of
ribulose-1,5-bisphosphate in the
Calvin cycle.
Related Topics