Chapter: Biochemistry: Photosynthesis
CO2 Fixation in Tropical Plants
CO2 Fixation in
Tropical Plants
In
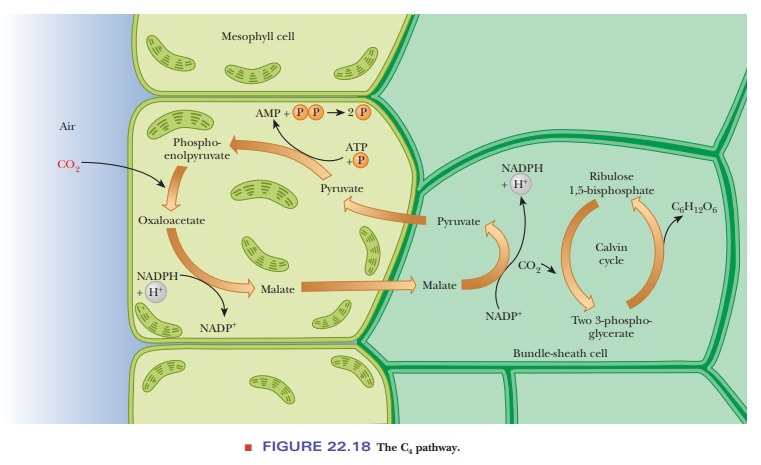
tropical plants, there is a C4 pathway (Figure 22.18), so named
because it involves four-carbon compounds. The operation of this pathway (also
called the Hatch–Slack pathway)
ultimately leads to the C3(based on 3-phosphoglycerate)pathway of
the Calvin cycle. (There are other C4 pathways, but this one is most
widely studied. Corn [maize] is an important example of a C4 plant,
and it is certainly not conÞned to the tropics.)
What is different about CO2 fixation in tropical plants?
When CO2
enters the leaf through pores in the outer cells, it reacts Þrst with
phosphoenolpyruvate to produce oxaloacetate and Pi in the mesophyll
cells of the leaf. Oxaloacetate is reduced to malate, with the concomitant
oxidation of NADPH. Malate is then transported to the bundle-sheath cells (the
next layer) through channels that connect the two kinds of cells.
In the
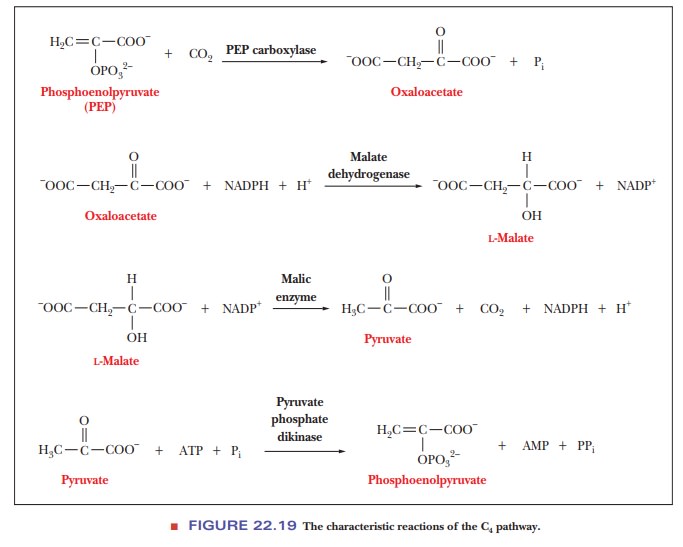
bundle-sheath cells, malate is decarboxylated to give pyruvate and CO2.
In the process, NADP+ is reduced to NADPH (Figure 22.19). The CO2
reacts with ribulose-1,5-bisphosphate
to enter the Calvin cycle. Pyruvate is transported back to the mesophyll cells,
where it is phosphorylated to phos-phoenolpyruvate, which can react with CO2
to start another round of the C4 pathway. When pyruvate is
phosphorylated, ATP is hydrolyzed to AMP and PPi. This situation
represents a loss of two high-energy phosphate bonds, equivalent to the use of
two ATP. Consequently, the C4 pathway requires two more ATP
equivalents than the Calvin cycle alone for each CO2 incorporated
into glucose. Even though more ATP is required for the C4 pathway
than for the Calvin cycle, there is abundant light to produce the extra ATP by
the light reaction of photosynthesis.
Note that the C4 pathway fixes CO2 in the mesophyll cells only to unfix it in the bundle-sheath cells, where CO2 then enters the C3 pathway. This observation raises the question of the advantage to tropical plants of using the C4 pathway.
The conventional wisdom on the subject focuses on the role of CO2,
but there is more to the situation than that. According to the conventional
view, the point of the C4 pathway is that it concentrates CO2
and, as a result, accelerates the process of photosynthesis. Leaves of tropical
plants have small pores to minimize water loss, and these small pores decrease
CO2 entry into the plant. Another point to consider is that the KM for CO2 of
phosphoenolpyruvate carboxylase is lower than that of rubisco, allowing the
outer mesophyll cells to fix CO2 at a lower concen-tration. This
also increases the concentration gradient of CO2 across the leaf and
facilitates the movement of CO2 into the leaf through the pores. In
tropical areas, where there is abundant light, the amount of CO2
available to plants controls the rate of photosynthesis.
The C4
pathway deals with the situation, allowing tropical plants to grow more quickly
and to produce more biomass per unit of leaf area than plants that use the C3
pathway. A more comprehensive view of the subject includes a consideration of
the role of oxygen and the process of photorespiration,
in which oxygen is used instead of CO2 during the reaction catalyzed
by rubisco.
Although
the actual biological role of photorespiration is not known, several points are
well established. The oxygenase activity appears to be an unavoid-able,
wasteful activity of rubisco. Photorespiration is a salvage pathway that saves
some of the carbon that would be lost because of the oxygenase activity of
rubisco. In fact, the photorespiration is essential to plants even though the
plant pays the price in loss of ATP and reducing power; mutations that affect
this pathway can be lethal. The principal substrate oxidized in
photorespiration is glycolate (Figure
22.20). The product of the oxidation reaction, which takes place in peroxisomes
of leaf cells, is glyoxylate.
(Photorespiration is localized in peroxisomes.) Glycolate arises ultimately
from the oxidative breakdown of ribulose-1,5-bisphosphate. The enzyme that catalyzes this reac-tion is
ribulose-1,5-bisphosphate
carboxylase/oxygenase, acting as an oxygen-ase (linked to O2) rather
than as the carboxylase (linked to CO2) that fixes CO2
into 3-phosphoglycerate.

When
levels of O2 are high compared with those of CO2,
ribulose-1,5-bisphosphate is
oxygenated to produce phosphoglycolate (which gives rise toglycolate) and
3-phosphoglycerate by photorespiration, rather than the two molecules of
3-phosphoglycerate that arise from the carboxylation reaction. This situation
occurs in C3 plants. In C4 plants, the small pores
decrease the entry not only of CO2 but also of O2 into
the leaves. The ratio of CO2 to O2 in the bundle-sheath
cells is relatively high as a result of the operation of the C4
pathway, favoring the carboxylation reaction. C4 plants have
successfully reduced the oxygenase activity by compartmentation and thus have
less need of photorespiration. This is an advantage in the hot climates in
which C4 plants are principally found.
Summary
In
tropical plants, four-carbon compounds are frequently involved in the pathway
of CO2 fixation.
This
alternative pathway facilitates the movement of CO2 into leaves and
prevents water loss.
Related Topics