Chapter: Biotechnology Applying the Genetic Revolution: Genomics and Gene Expression
Gaps Remain in the Human Genome
GAPS
REMAIN IN THE HUMAN GENOME
Although the sequence of the
genome is considered complete, there are still gaps. The gaps fall in highly
condensed regions of highly repetitive heterochromatin
that is difficult to sequence. Three features characterize heterochromatin: hypoacetylation
(i.e., lack of acetyl groups on the histones); methylation of histone H3 on a
specific lysine; and methylation on CpG or CpNpG sequence motifs.
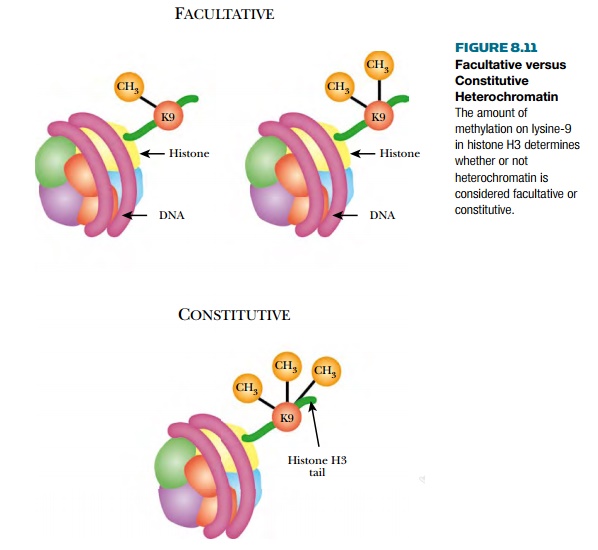
Heterochromatin is not transcribed and comes in two forms, facultative heterochromatin and
constitutive heterochromatin (Fig. 8.11). The constitutive form is found around the centromeres and
telomeres of the chromosome and does not change from one generation to the
next.
Facultative heterochromatin,
on the other hand, is found in other regions of the chromosomes, and its
presence is cell-specific. Once a specific region of a chromosome becomes
heterochromatin, all of its descendent cells will maintain this pattern.
However, neighboring cells may not have heterochromatin in the same regions,
leading to position effect variegation (PEV). The theory is that
heterochromatin forms in defense against invading retrotransposons and viruses,
and also due to gene silencing during RNAi. When a gene is near a region of
heterochromatin, the gene is no longer transcribed and becomes dormant.
The amount of methylation on
lysine-9 in histone H3 determines whether or not heterochromatin is considered
facultative or constitutive.
Related Topics