Chapter: Psychology: Motivation and Emotion
The Diversity of Motives
THE DIVERSITY
OF
MOTIVES
Some
of the wide range of motives we have discussed are well described in drive
terms. Other motives seem to have both a prominent avoidance (drive-reduction)
component and an approach component. As this list of motives grows, however,
one pressing ques-tion is how to organize the many motives that energize and
direct our behavior.
Maslow and the Hierarchy of Needs
One
approach was suggested by Maslow, who described a hierarchy of needs in which the lower-order physiological needs are
at the bottom of the hierarchy, safety needs are further up, the need for
belonging is still higher, and the desire for esteem is higher
yet. Higher still, toward the
top of the
hierarchy, is the striving
for self- actualization—the desire to realize oneself to the fullest
(Figure 12.25).
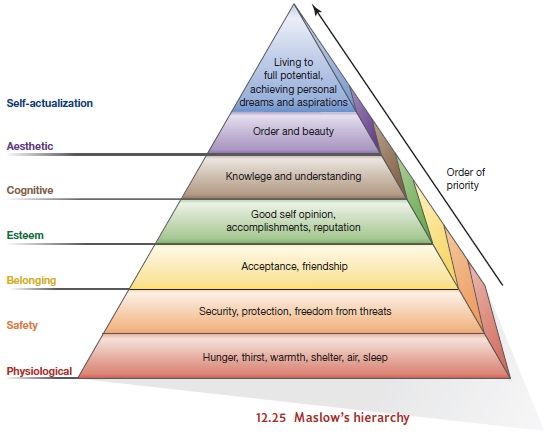
Maslow
believed that people will strive for higher-order needs (e.g., self-esteem or
artis- tic achievement) only when lower-order needs (e.g., hunger) are
satisfied. By and large this is plausible enough; the urge to write poetry gen-
erally takes a back seat if one has not eaten for days. But
as Maslow pointed
out, there are exceptions. Some
artists starve rather
than give up their
poetry or their
painting, and some martyrs
proclaim their faith
regardless of pain or
suffering. But to
the extent that Maslow’s
assumption holds, a motive
toward the top of his hierarchy—such as
the drive toward self-actualization—will become
pri- mary only when all other needs beneath it are satisfied.
The Twin Masters: Pain and Pleasure
A
second approach to organizing our many motives, rather than listing them
hierarchi-cally as Maslow did, is to dig deeper and look for a few common
principles that under-lie our diverse motives. This approach leads us to
consider the twin masters of pain and pleasure which may be said to govern much
of our behavior.
THE AVOIDANCE OF PAIN
What
do being too hot (or cold), being famished, being frightened, and being
sexually frustrated have in common? We sometimes speak of each as involving its
own type of pain, although this usage
of the word might seem metaphorical. After all, in discussing a specific
sensation typically associated with tissue injury or irritation. It turns out,
though, that there is no metaphor here, and various states of discomfort do
involve mechanisms overlapping with those associated with, say, stepping on a
nail or bumping your head.
As
we saw, pain is a “general purpose” signal that all is not well. There are
multiple pain pathways, including a fast pain pathway that detects localized
pain and relays this information quickly via the thalamus to the cortex using
thick, myelinated fibers, as well as a slow pain pathway that carries less
localized sensations of burning or aching via the thalamus to subcortical brain
structures such as the amygdala using thinner unmyelinated fibers. These
pathways differ in important ways, but they both carry information that allows
us to distinguish different types of painful stimulation, and an awareness of
these differences is important, because this is what allows us to know what
steps to take to decrease the pain we are feeling.
At
the same time, researchers are beginning to appreciate that many different
types of stimulation activate a common brain network, referred to as the pain matrix. The pain matrix consists
of a distributed set of regions including the thalamus and the anterior cingulate
cortex. The physical pain of accidentally touching a hot stove acti-vates this
matrix, but so does the psychological pain that comes with being socially
snubbed, and this is what tells us that these various forms of discomfort
really do have elements in common—both functionally and neurally. Figure 12.26
shows the similar brain activations associated with thermal stimulation (Panel
A) as well as social exclu-sion (Panel B).
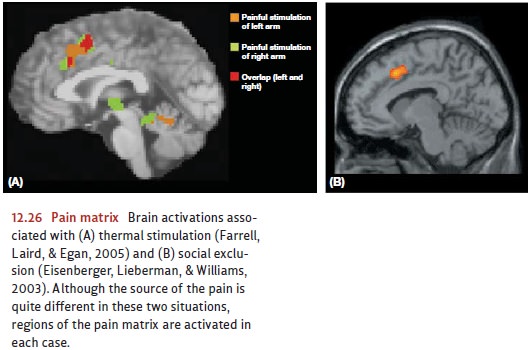
What’s
important here is to appreciate that pain in all of its forms has both a
specific and a general motivational role. Some pain signals provide specific
information about what is happening (there’s a burning sensation in my right
hand) and motivate a specific response (I need to move my hand away from the
stove). Other pain signals alert us to different, broader kinds of problems,
and suggest different types of responses, such as a departure from a socially
awkward situation.
This
overlap among very different types of “pain” is important for another reason.
It provides a “common currency” in which to express a wide range of undesirable
states. The function of this common currency becomes clear when we realize that
many motives typically operate at a time, ranging from tight shoes to thirst to
fatigue. In such settings, the generic aspect of pain allows us to decide which
of these quite different states to attend to now and which can be handled
later.
THE PURSUIT OF PLEASURE
Efforts
to avoid or reduce painful states of tension govern many of our behaviors.
However, as Maslow pointed out, reducing these states is not our only motive.
We also have positive goals, and seek out pleasurable states.
Such
states involve incentives, or positive goals that we seek to obtain. We
therefore do not just feel “pushed” by an internal state of tension. Instead,
the incentive seems to “pull” us in a certain direction. As we have seen,
though, these incentives come in different types. Some are an inherent part of
the activity or object to which we are drawn (e.g., playing the guitar because the
experience itself is fun). In such
cases, the activity or object is said to be intrinsically rewarding. Other incentives, by contrast, are not an
integral part of the activity or object to which we are drawn, but are instead
arti-ficially associated with it (e.g., getting paid for painting a picture).
In this case, we say that the activity or object is extrinsically rewarding.
Historically,
research on incentives has focused on external determinants of behavior— the
specific rewards available to us and their influence on our behavior. It is
becoming clear, however, that internal states of pleasure have a role in
supporting incentives that parallels the role of the internal states of pain in
supporting drives. This realization has directed researchers’ attention to
examining two different aspects of our responses to rewarding stimuli, namely,
wanting and liking.
Wanting refers to the organism’s
motivation to obtain a reward and is measured bythe amount of effort the
organism will exert to obtain it. Because wanting is defined behaviorally, it
can be assessed in any organism whose behavior we can measure. With rats, we
can ask how many times they will press the bar to dispense a food pellet. In a
similar fashion, we can see how hard people will work to obtain a reward: If
you want brownies, do you want them badly enough to go through the trouble of
making them? And if you don’t have the ingredients, do you want brownies badly
enough to first go shopping and then make them?
In
contrast to wanting, liking refers
to the pleasure that follows receiving the reward, like the pleasure deriving
from the luscious taste of the warm brownies. We all know the sensation of
liking, but unlike wanting, liking is difficult to define behaviorally.
However, we can explore the neural basis of this experience. Research on this
point dates back more than half a century to work by Olds and Milner, who
electrically stimulated rats’ brains in different regions. They found that
stimulation in some regions led rats to engage in behaviors such as returning
repeatedly to the location at which they were ini-tially stimulated, seemingly
in an attempt to repeat the stimulation. To understand this phenomenon, Olds
and Milner made the brain stimulation contingent upon bar presses, and found
that rats would bar press at very high rates for long periods of time, and to
the exclusion of other activities, in order to obtain this stimulation (Figure
12.27). One par-ticularly important spot for this effect—in rats and
humans—appears to be the medial forebrain bundle, a nerve tract consisting of
neurons with cell bodies in the midbrain which synapse in the nucleus accumbens (in the striatum).

Apparently,
stimulation in this brain area is strongly rewarding—but why? Is this area involved in wanting
or in liking? Modern neuroimaging
research suggests an answer, and indicates that different brain regions are
engaged by reward anticipation (wanting) and reward delivery (liking) (Schultz,
Tremblay, & Hollerman, 2000). In particular, regions in the frontal cortex
are activated by liking, whereas the nucleus accumbens is especially sensitive
to wanting. In one of the first studies to demonstrate this dissociation in
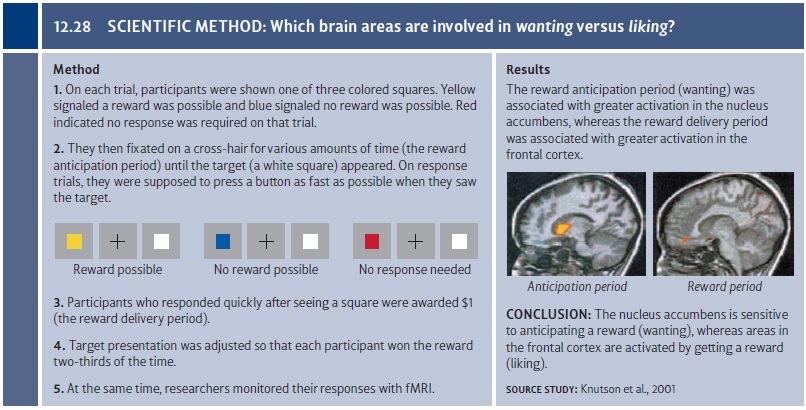
humans, Knutson and colleagues used fMRI during a task in which participants
were shown various types of cues and were asked to respond to a target
(Knutson, Fong, Adams, Varner, & Hommer, 2001). If participants responded
quickly enough, they were able to win one dollar, and target presentation was
adjusted for each individual so that each participant succeeded approximately
two-thirds of the time. As shown in Figure 12.28, the period of anticipation and
wanting was associated with greater activation in the nucleus accumbens,
whereas the period of liking the reward was associated with activation in the
frontal cortex. These activation patterns suggest that the processes of wanting and liking have separate bases in the brain.
Additional
evidence that wanting and liking depend on separable brain systems comes from
studies of the neurotransmitters that underlie reward processing. When
animals
are trained to bar press for a reward, their neurons release dopamine into the
nucleus accumbens neurons during anticipation but not during receipt of reward
(Phillips et al., 2003). When dopamine antagonists are introduced, which
interfere with the effects of dopamine, the animals will continue to engage in
incentive-driven behavior for a time, but they will not work to receive rewards
that are not present (Berridge & Robinson, 2003). These and other findings
suggest that dopamine release plays a central role in wanting. Complementary
studies have examined the neurotrans-mitters that are responsible for liking.
These studies suggest that endorphins are released into the nucleus accumbens
when rewards are delivered. When endorphin antagonists are administered (in humans),
these appear to diminish the subjective pleasure associated with consuming the
rewards (Yeomans & Gray, 1996).
Related Topics