Chapter: Microbiology
Microbial nutrition and growth
MICROBIAL NUTRITION AND GROWTH
‘An army marches on its stomach’ said Napolean Bonapart. This indicates that food is important for any living or-ganism and so also for microbes. Food is any substrate that can be metabolized to provide assimilable material or energy for the cell. Plants synthesize their own food requirements through photosyn-thesis. Animals ingest the presynthesized food from plants or by devouring other animals. All living organisms, from micro to macroorganisms require nutrients for growth and normal function-ing. Animals ingest the food and digest them in their digestive system (Holozoic nutrition) to simpler nutrients which are absorbed by cells for synthesis of all cellular material and derive energy. Plants absorb the nutrients from soil solution (Holophytic nutri-tion) released by mineralisation of organic matter and grow. Mi-croorganisms particularly fungi derive their nutrients from the ex-tra corporeal digestion by secreting extracellular enzymes. The nutrients are absorbed and cellular materials are synthesized.
All organisms exhibit two universal requirements viz., water and elements. All organisms require energy which they de-rive from the chemical compounds or radiant energy like light. The elemental components are carbon, nitrogen, phosphorus, sul-phur and potassium besides hydrogen and oxygen major ones for synthesizing cellular components. Metal ions like K, Ca, Mg and Fe are required for normal growth. Other metal ions like Zn, Cu, Mn, Mo , Ni, B, Co are often required in low quantities hence known as trace elements. Fe, Mg, Zn, Mo, Mn and Cu are cofac-tors/coenzymes or prosthetic group of various enzymes. Most bac-teria do not require Na but certain marine bacteria, cyanobacteria and photosynthetic bacteria require it. Red extreme halophiles can- not grow with less than 12 to 15% NaCl which is required to maintain the integrity of cell walls and for the stability and activity of certain enzymes. Silicon is required for the growth of diatoms.
Vitamins and vitamin like compounds are also present in living cells. These function either as coenzymes or as building blocks of coenzymes. Some bacteria synthesize their entire requirements of vitamins but some cannot grow unless supplied from external source.
Microorganisms are divided into several types based on the energy source or electron source and carbon assimilation. Those derive energy from the oxidation of chemical compounds are known as ‘chemotrophs’ and others utilizing radiant energy like light are known as ‘phototrophs’. Electrons are required for me-tabolism and based on the source from which bacteria derive elec-tron they are grouped. Some organisms use reduced inorganic com-pounds as electron donors and are termed as ‘lithotrophs’ literally meaning rock eating. Others use organic compounds are termed as ‘organotrophs’. Those organisms that derive energy from the chemical compounds (Chemotrophs) and uses inorganic com-pounds as e- donors (lithotrophs) are known as chemolithotrophs. Those that derive energy from light (phototrophs) and e- from in-organic compounds are photolithotrophs. Similarly those chemotrophs that use organic compounds, as e- donors are chemoorganotrophs and the phototrophs that utilize organic compounds as e- donors are photoorganotrophs
Chromatium okenii, a photosynthetic bacterium, uses radi-ant energy and H2S as electron donor oxidizing it to elemental sulphur. Some phototrophs use organic compounds such as fatty acids and alcohols as electron donors and hence called photoorganotrophs.
Rhodospirillum rubrum another phototrophic bacterium uti-lizes succinate as e- donor converting it to fumarate. A phototrophic bacterium can grow as chemotroph. In the anoxygenic environ-ment (absence of O2) this bacterium grow as photoor but in the presence of oxygen and dark (absence of light) it grows as a chemoorganotroph. Among the chemotrophs some utilize inorganic
compounds like NH4 as e- donors and hence called chemolithotrophs. Nitrosomonas use ammonia for electrons and derive energy by oxidizing ammonia to nitrite. Certain chemotrophs use organic compounds like sugars and amino acids as e- donor
and are called chemoorganotrophs. Some of the chemotrophs can grow either as chemolithotrophs or chemoorganotrophs. Pseudomonas pseudofulvacan use glucose an organic compound (chemoorganotrophs) or inorganic compound H2 as e- source (chemolithotrophs)
Autotrophs and Heterotrophs
Based on the source of carbon microorganisms are grouped as autotrophs and heterotrophs. Some can use CO2 as their sole source of carbon like plants and algae are termed as autotrophs. Others like some bacteria, fungi and actinomycetes utilize preformed organic compounds as carbon source and hence called heterotrophs.
Most organisms that involve in decomposition of organic matter in soil are heterotrophs Fungi are saprophytic and depend on dead organic matter. Some fungi are parasitic on living plants and animals. The saprophytic and parasitic organisms are heterotrophs. Such of these heterotrophs that have elaborate requirements of specific nutrients like vitamins and growth promoting substances are called fastidious heterotrophs as they are not easily pleased or satisfied by ordinary nutrients available in nature.
The source of carbon for microbes is CO2 or carbohydrates. Autotrophs derive their entire requirement of C from CO2 while heterotrophs derive the carbon chiefly from carbohydrate. In nature, cellulose, hemicelluloses, starch, pectin, lignin etc serve as
carbon sources. Amino acid, purine and pyrimidine bases, protein serve as a source of nitrogen. Phosphorus is obtained from the nucleotides, phytin etc. For cultivation of microorganisms in labo-ratory, media containing monosaccharides like glucose and disac-charides like sucrose are used as C sources. Peptone, Tryptone, inorganic salts like ammonium salts, potassium nitrate serve as nitrogen sources. Potassium dihydrogen phosphate and dipotas-sium hydrogen phosphate are commonly employed to serve as sources of phosphorus and also as a buffering agent.
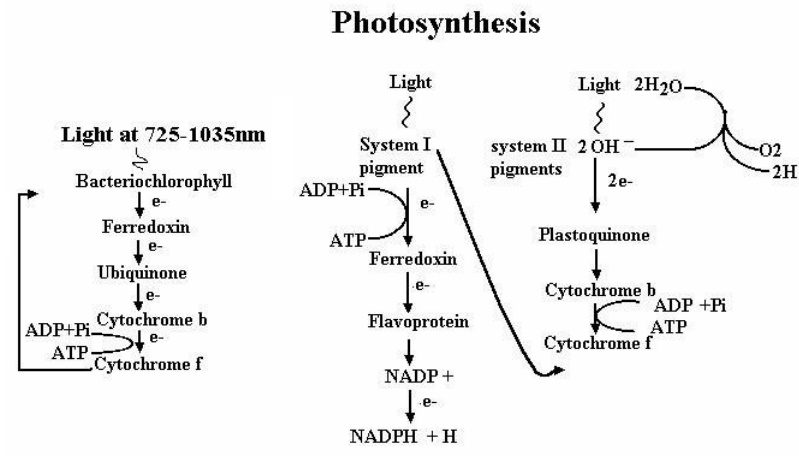
Autotrophic bacteria have the simplest nutritional require-ments as they can grow and reproduce in a mixture of inorganic compounds. They also possess an elaborate capacity to synthesize the carbohydrate, proteins, lipids, nucleic acids, vitamins and other complex substances of living cells. Photosynthesis is a normal autotrophic way of life and this occurs in plants, algae, photosyn-thetic bacteria and cyanobacteria. In this process, CO2 is reduced and converted to carbohydrate utilizing light. However, photosyn-thesis of plants, algae and cyanobacteria perform oxygen evolving photosynthesis by absorbing the reducing power from the pho-tolysis of water. On the other hand, photosynthetic bacteria green and purple bacteria obtain the reducing power from a compound similar to water (H20) viz., H2S i.e. available in anoxygenic envi-ronment. The pigments and the light absorption also differ in these organisms.
Over all reaction of photosynthesis is,
2H20 + CO2−−> (C2H0) x + O2 + H20 (Plants, algae, cyanobacteria)
2H2S+ CO2 −−> (C2H0) x + 2S + H20 (Photoautotroph bacteria)
Inorganic compounds like H2, H2S203 or the organic com-pounds lactate, succinate can be the source of reducing power in-stead of H2S.
Growth
Living organisms grow and reproduce. The growth indicates that an organism is in active metabolism. In plants and animals one see the increase in height or size. In a butterfly, a small larva hatching from egg grows in size, moults, pupates and become an adult butterfly through metamorphosis. Growth in a common use refers to increase in size but with microorganisms particularly with bacteria, this term refers to changes in total population rather than increase in size or mass of an individual organism. With fungi linear growth of hyphae and radial growth of colony is observed for growth on solid media but a biomass or mycelial dry weight on liquid media. In unicellular fungi like yeast that reproduce by fis-sion or budding the population change is considered as growth.
The change in population in bacteria chiefly involves trans-verse binary fission in most of the bacteria while budding is ob-served in Hyphomicrobium. In actinomycetes, fragmentation of hyphae and sporulation results in population change. In yeasts, budding and fission are observed that this depends upon the spe-cies. In fungi, growth fragmentation asexual and sexual spores serve as propagates for population increase.
The transverse binary fission, an asexual reproductive pro-cess is the most common in the growth cycle of bacterial popula-tion. A single cell divides after developing a transverse septum (cross wall) and continues to grow by continuous dividing with-out cell death till it is subjected to stress.
A cell dividing by binary fission is immortal unless subjected to stress by nutrient depletion or environmental stress. Therefore a single bacterium continuously divides. One cell divides providing two cells and two cells divide giving four and so on. Therefore the population increases by geometric progression.

When a single bacterium is incubated into the liquid me-dium in flask and incubated, bacterium divides by fission and at-tains a period of rapid growth in which cells multiply at an expo-nential rate. If the logarithmic number of bacterium versus time is plotted a growth curve is obtained with different phases of growth.
Soon after transfer of an inoculum to a new medium, cells do not immediately multiply and the population remains un-changed. The cells however increase in size synthesizing newer protoplasm and enzyme necessary to the newer environment. The organisms are metabolizing but require more for adjustments to the physical environment around each cell and hence there is a lag for cell division (lag phase).
At the end of lag phase cells divide and there is a gradual increase in the population. When all the cells complete their lag, there is division at regular intervals. The cells divide steadily at a constant rate in the logarithmic or exponential phase and when log number of cells are plotted against time there is a straight line. The population in this phase is almost uniform in chemical composi-tion, metabolic activity and physiological characteristics.
Generation time is the time required for the population to double and this can be determined by the number of generation that occurs at a particular time interval. Not all bacteria have the same generation time. It varies from 15 – 20 minutes for chia coli to many hours in others and is also dependent upon thenutrients and physical conditions of the environment. With the growth of the bacterium, there will be a depletion of nutrients. At high concentration of nutrients a small change may not cause sig-nificant effect but at low concentration the growth rate decreases significantly.
At the end of the exponential phase growth rate decreases due to exhaustion of some nutrients or due to production of toxic products during growth. The population remains constant due to complete cessation of division or reproduction rate equals to death rate.
The stationary phase is followed by Decline or Death phase as bacteria divide faster than the new cells produced. The deple-tion of nutrients, accumulation of solubilising products like acids.
The number of viable cells decreases exponentially. G-ve Cocci divide faster but others divide slowly but viable cells may persist for minutes or even years.
Measurement of growth
Growth refers to the magnitude of the population in bacteria. The growth can be measured quantitatively (1) cell count (2) cell mass and (3) cell activity. Cell count shall be made directly by microscopy or using an electronic particle counter. It can also be made indirectly by colony count after serially diluting the sample. Cell mass can be determined directly by weighing a known vol-ume of sample culture broth or by measuring the cell nitrogen. It can also be determined indirectly by finding cell activity, which can be measured by the degree of biochemical activity to the size of population.
Petroff – Hausen counting chamber is used for direct micro-scopic count. It is a slide accurately ruled into squares of 1/400 mm2 area over when a cover slip rests at 1/50mm above. This gives a volume of 1/20000mm3 over one square. The liquid can be placed in the chamber left unstained and counted using a phase contrast microscopy. If 5 cells are present in one square there will be 5 X 20,000,000 or 108 cells/ml. This method is rapid requires simple equipment. Morphology of cells can be simultaneously obtained but difference of viable or dead cells cannot be made.
In electronic particle counter a bacterial suspension is passed through a tiny orifice of 10-30 μm diameter that connects the two comparts of counter containing an electrically conductive solu-tion. The electrical resonance between the two compartments in-crease momentary when each bacterial cell passes in the orifice creating an electrical signal. The signals are automatically counted. This method is rapid but requires sophisticated electronic equip-ment.
In plate count method a known volume of bacterial suspen-sion diluted serially if population is dense, is poured in petridishes and molten agar medium is added and mixed thoroughly. The cells trapped immediately in the gelled medium develop into a colony. The colonies are counted by illumination from below. The num-ber of colonies is multiplied by the dilution factor to detect the population and expressed as number per ml. Only those cells that grow in the medium that show up and get counted under the con-ditions in which the plates are incubated. If aggregate of cells are formed as in cocci in clusters, chains or pairs the resulting counts will be lower than the individual cells. In such cases counts are referred to as colony – forming units per ml than number per ml. The plate count is used normally for estimation of bacterial popu-lation in milk, waste foods, soil and many other materials.
Mernbrane filter count is used to determine the bacterial numbers in large sample containing very small number of viable cells. Bacteria in large volume of air or wastes shall be collected by filtering in the membrane. The membrane with filtered bacteria is placed on plate containing a pad saturated with medium and incubated. The organisms grow into a colony and counted. Special medium and dyes can be used to detect certain types of organ-isms.
The bacteria in suspension absorb and scatter light passing through the cell similar to water droplets in fogs absorbing and scattering of light. Because of this phenomenon, a culture of more than 107 or 108 cells per ml appears turbid to the naked eye.
The cell growth can also be measured by the nitrogen con-tent that forms the process when is a major constituent of cell. Cells are harvested, washed free of medium and nitrogen is analysed by standard method.
The quantitative measurement of a mass of cells is made by the dry weight determination. Very dense suspension of cells can be washed free of extraneous matter and weighed. In cells accu-mulating β –hydroxy butyrate cell mass may increase without corre-sponding increase in cell growth.
The measurement of acid or any other product of metabolism shall be measured to assess growth.
In case of yeasts, dry weight determination and nitrogen estimation can be done as a measure of growth. In mycelial fungi, mycelial dry weights are determined by filtering the mycelial mat in a previously weighed filter paper drying it in oven at 105oC for 24 hours and weighing it. The mycelial weight is determined by subtracting the weight of filter paper. In agar medium, the linear growth / nodal growth of fungi shall be measured.
Related Topics