Chapter: Essential Microbiology: Microbial Nutrition and Cultivation
How do Microbial nutrients get into the microbial cell?
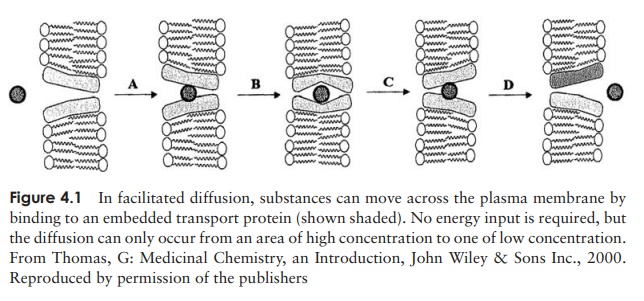
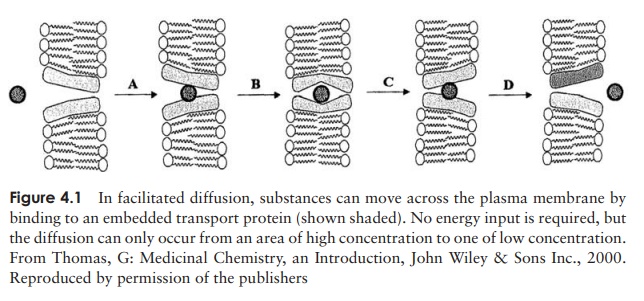
How do nutrients get into the microbial cell?
Having found a source of a given nutrient, a microorganism must:
· have some means of taking it up from the environment
· possess the appropriate enzyme systems to utilise it.
The plasma membrane represents a selective barrier, allowing into the cell only those substances it is able to utilise. This selectivity is due in large part to the hydrophobic nature of the lipid bilayer. A substance can be transported across the cell membrane in one of three ways, known as simple diffusion, facilitated diffusion and active transport.
In simple diffusion, small molecules move across the membrane in response to a concentration gradient (from high to low), until concentrations on either side of the membrane are in equilibrium. The ability to do this depends on being small (H2O, Na+ , Cl− ) or soluble in the lipid component of the membrane (non-polar gases such as O2 and CO2).
Larger polar molecules such as glucose and amino acids are unable to enter the cell unless assisted by membrane-spanning transport proteins by the process of facilitated diffusion (Figure 4.1). Like enzymes, these proteins are specific for a single/small number of related solutes; another parallel is that they too can become saturated by too much ‘substrate’. As with simple diffusion, there is no expenditure of cellular energy, and an inward concentration gradient is required. The transported substance tends to be metabolised rapidly once inside the cell, thus maintaining the concentration gradient from outside to inside.
Diffusion is only an effective method of internalising substances when their concen-trations are greater outside the cell than inside. Generally, however, microorganisms find themselves in very dilute environments; hence the concentration gradient runs in the other direction, and diffusion into the cell is not possible. Active transport enables the cell to overcome this unfavourable gradient. Here, regardless of the direction of the gradient, transport takes place in one direction only, into the cell. Energy, derived from
hydrolysis of adenosine triphosphate is required to achieve this, and again specific transmembrane proteins are involved. They bind the solute molecule with high affinity outside of the cell, and then undergo a conformational change that causes them to be released into the interior. Procaryotic cells can carry out a specialised form of active transport called group translocation, whereby the solute is chemically modi-fied as it crosses the membrane, preventing its escape. A well-studied example of this is the phosphorylation of glucose in E. coli by the phosphotransferase system. Glucose present in very low concentrations outside the cell can be concentrated within it by this mechanism. Glucose is unable to pass back across the membrane in its phosphorylated form (glucose-6-phosphate), however it can be utilised in metabolic pathways in this form.
Often it may be necessary to employ extracellular enzymes to break down large molecules before any of these mechanisms can be used to transport nutrients into the cell.
Related Topics