Chapter: Essential Microbiology: Cell Structure and Organisation
The procaryotic cell and its structure
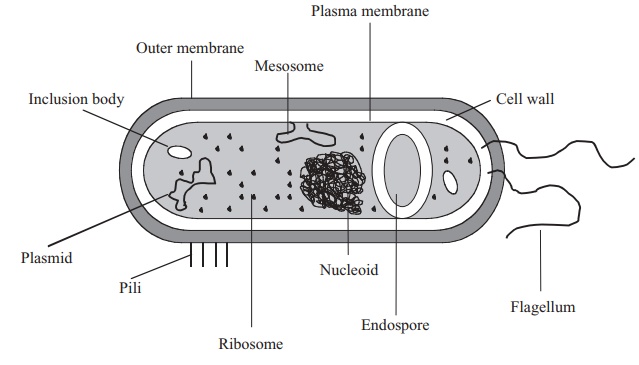
The procaryotic cell
Bacteria are much smaller than eucaryotic cells; most fall into a size range of about 1–5µm, although some may be larger than this. Some of the smallest bacteria, such as the mycoplasma measure less than 1µm, and are too small to be resolved clearly by anordinary light microscope.
Because of their extremely small size, it was only with the advent of the electron microscope that we were able to learn about the detailed structure of bacterial cells. Usingthe light microscope however, it is possible to recognize differences in the shape and arrangement of bacteria. Al-though a good deal of variation is possible, most have one of three basic shapes (Figure 3.2):
· rod shaped (bacillus)
· spherical (coccus)
· curved: these range from comma-shaped (vibrio) to corkscrew-shaped (spirochaete)
All these shapes confer certain advantages to their owners; rods, with a large surface area are better able to take up nutrients from the environment, while the cocci are less prone to drying out. The spiral forms are usually motile; their shape aids their movement through an aqueous medium.
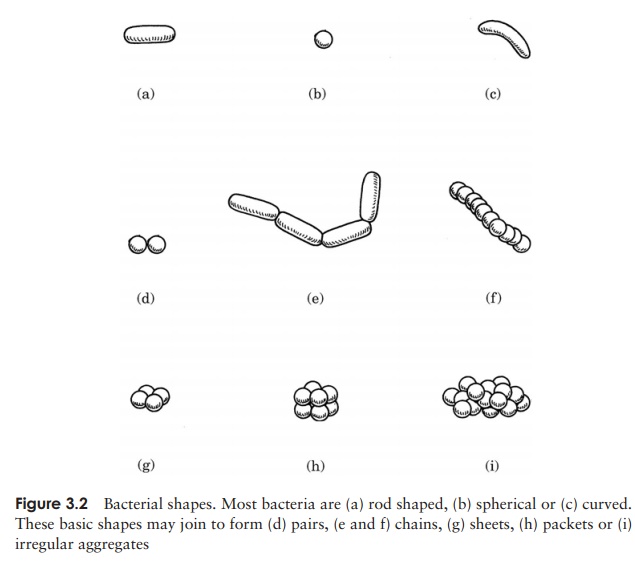
As well as these characteristic cell shapes, bacteria may also be found grouped to-gether in particular formations. When they divide, they may remain attached to one another, and the shape the groups of cells assume reflects the way the cell divides. Cocci, for example, are frequently found as chains of cells, a reflection of repeated division in one plane (Figure 3.2(f)). Other cocci may form regular sheets or packets of cells, as a result of division in two or three planes. Yet others, such as the staphylococci, divide in several planes, producing the irregular and characteristic ‘bunch of grapes’ appear-ance. Rod-shaped bacteria only divide in a single plane and may therefore be found in chains, while spiral forms also divide in one plane, but tend not to stick together. Blue–greens form filaments; these are regarded as truly multicellular rather than as a loose association of individuals.
Procaryotic cell structure
When compared with the profusion of elaborate organelles encountered inside a typical eucaryotic cell, the interior of a typical bacterium looks rather empty. The only internal structural features are:
· a bacterial chromosome or nucleoid, comprising a closed loop of double stranded, supercoiled DNA. In addition, there may be additional DNA in the form of aplasmid
· thousands of granular ribosomes
· a variety of granular inclusions associated with nutrient storage.
All of these are contained in a thick aqueous soup of carbohydrates, proteins, lipids and inorganic salts known as the cytoplasm, which is surrounded by a plasma membrane. This in turn is wrapped in a cell wall, whose rigidity gives the bacterial cell its characteristic shape. Depending on the type of bacterium, there may be a further surrounding layer such as a capsule or slime layer and/or structures external to the cell associated with motility (flagella) or attachment (pili/fimbriae). Figure 3.3 shows these features in a generalised bacterial cell. In the following pages we shall examine these features in a little more detail, noting how each has a crucial roleto play in the survival or reproduction of the cell.
Genetic material
Although it occupies a well defined area within the cell, the genetic material of procaryotes is not present as a true nucleus, as it lacks a surrounding nuclear membrane (c.f. the eucaryotic nucleus, Figure 3.12). The nucleoid or bacterial chromosome comprises a closed circle of dou-ble stranded DNA, many times the length of the cell and highly folded and compacted. (The common laboratory
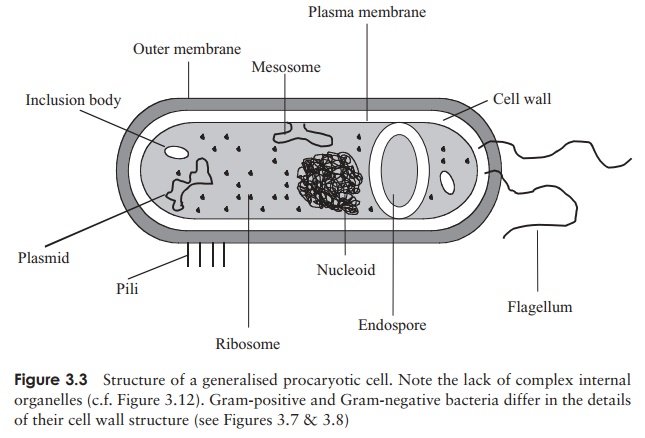
Bacterium Escherichia coli is around 3–4 µm in length, but contains a DNA molecule some 1400 µm in length!) The DNA may be associated with certain bacterial pro-teins, but these are not the same as the histones found in eucaryotic chromosomes. Some bacteria contain ad-ditional DNA in the form of small, self-replicating ex-trachromosomal elements called plasmids. These do not carry any genes essential for growth and reproduction, and thus the cell may survive without them. They can be very important however, as they may include genes encoding toxins or resistance to antibiotics, and can be passed from cell to cell.
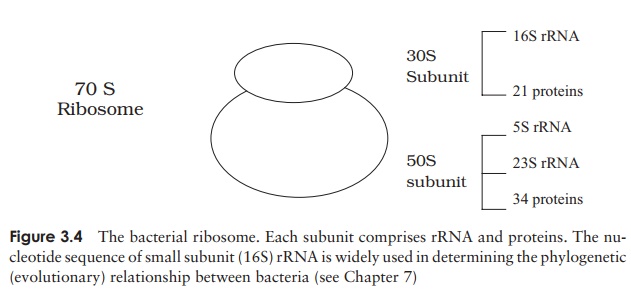
Ribosomes
Apart from the nucleoid, the principal internal structures of procaryotic cells are the ribosomes. These are the site of protein synthesis, and there may be many thousands of these in an active cell, lending a speckled appearance to the cytoplasm. Ribosomes are composed of a complex of protein and RNA, and are the site of protein synthesis in the cell.
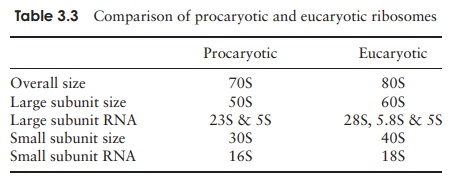
Although they carry out a similar function, the ribosomes of procaryotic cells are smaller and lighter than their eucaryotic counterparts. Ribosomes are mea-sured inSvedberg units (S), a function of their size and shape, and determined by their rate of sedimentation in a centrifuge; procaryotic ribosomes are 70S, while those of eucaryotes are 80S. Some types of antibiotic exploit this difference by
Inclusion bodies
Within the cytoplasm of certain bacteria may be found granular structures known as inclusion bodies. These act as food reserves, and may contain organic compounds such as starch, glycogen or lipid. In addition, sulphur and polyphosphate can be stored as inclusion bodies, the latter being known as volutin or metachromatic granules. Two special types of inclusion body are worthy of mention. Magnetosomes, which contain a form of iron oxide, help some types of bacteria to orientate themselves downwards into favourable conditions, whilst gas vacuoles maintain bouyancy of the cell in blue greens and some halobacteria.
EndosporesCertain bacteria such as Bacillus and Clostridium pro- duce endospores. They are dormant forms of the cell that are highly resistant to extremes of temperature, Ph and other environmental factors, and germinate into new bacterial cells when conditions become more favourable.
The spore’s resistance is due to the thick coat that surrounds it.
The plasma membrane
The cytoplasm and its contents are surrounded by a plasma membrane, which can be thought of as a bilayer of phospholipid arranged like a sandwich, together with associated proteins (Figure 3.5). The function of the plasma membrane is to keep the contents in, while at the same time allowing the selective passage of certain substances in and out of the cell (it is a semipermeable membrane).
Phospholipids comprise a compact, hydrophilic (= water-loving) head and a long hydrophobic tail region (Figure 2.27); this results in a highly ordered structure when the membrane is surrounded by water. The tails ‘hide’ from the water to form the inside of the membrane, while the heads project outwards. Also included in the membrane are a variety of proteins; these may pass right through the bilayer or be associated with the inner (cytoplasmic) or outer surface only. These proteins may play structural or functional roles in the life of the cell. Many enzymes associated with the metabolism of nutrients and the production of energy are associated with the plasma membrane in procaryotes. As we will see later, this is fundamentally different from
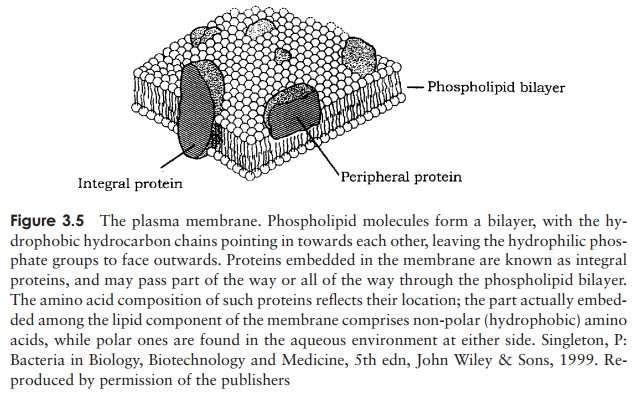
eucaryotic cells, where these reactions are carried out on specialised internal organelles. Proteins involved in the active transport of nutrients are also to be found associated with the plasma membrane. The model of membrane structure as depicted in Figure 3.5 must not be thought of as static; in the widely accepted fluid mosaic model, the lipid is seen as a fluid state, in which proteins float around, rather like icebergs in an ocean.
The majority of bacterial membranes do not contain sterols (c.f. eucaryotes: see below), however many do contain molecules called hopanoids that are derive from the same precursors. Like sterols, they are thought to assist in maintaining membrane stability. A comparison of the lipid components of plasma membranes reveals a distinct difference between members of the Archaea and the Bacteria.
The bacterial cell wall
Bacteria have a thick, rigid cell wall, which maintains the integrity of the cell, and determines its characteristic shape. Since the cytoplasm of bacteria contains high concentrations of dissolved substances, they generallylive in a hypotonic environment (i.e. one that is more di-lute than their own cytoplasm). There is therefore a nat-ural tendency for water to flow into the cell, and withoutcell wall the cell would fill and burst (you can demon- strate this by using enzymes to strip off the cell wall, leaving the naked protoplast).
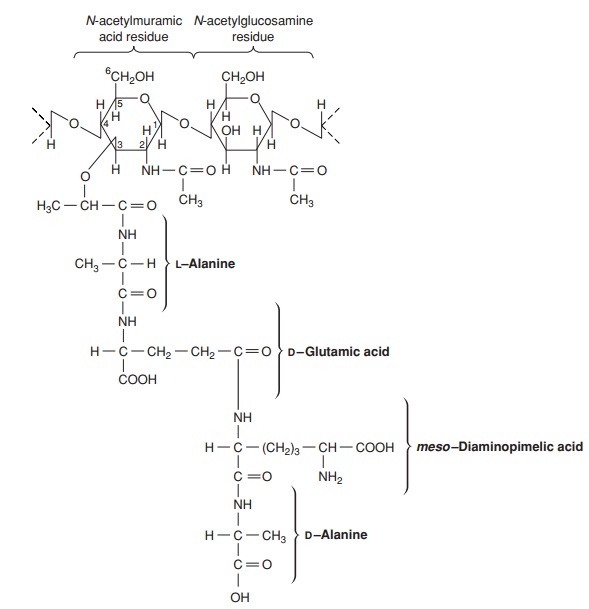
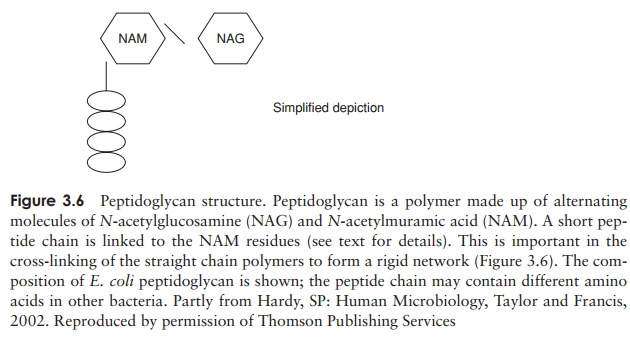
The major component of the cell wall, which is re-sponsible for its rigidity, is a substance unique to bacte-ria, called peptidoglycan (murein). This is a high molecular weight polymer whose basic subunit is made up ofthree parts: N-acetylglucosamine, N-acetylmuramic acid and a short peptide chain (Fig-ure 3.6). The latter comprises the amino acids L-alanine, D-alanine, D-glutamic acid and either L-lysine or diaminopimelic acid (DAP). DAP is a rare amino acid, only found in the cell walls of procaryotes. Note that some of the amino acids of peptidoglycan are found in the D-configuration. This is contrary to the situation in proteins, as you may recall, and confers protection against proteases specifically directed against L-amino acids.
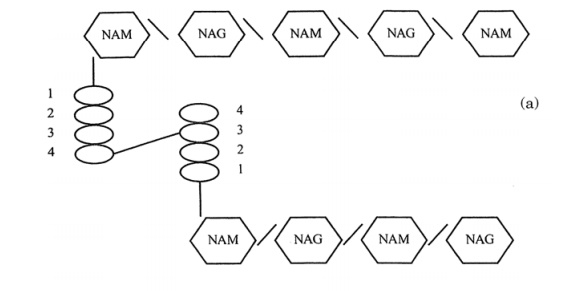
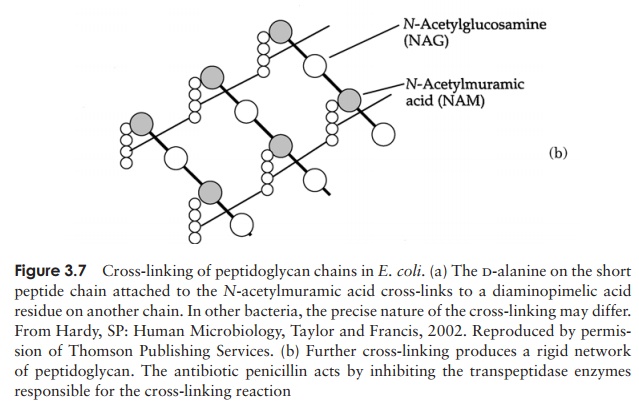
Precursor molecules for peptidoglycan are synthesised inside the cell, and transported across the plasma membrane by a carrier called bactoprenol phosphate before being in-corporated into the cell wall structure. Enzymes called transpeptidases then covalently bond the tetrapeptide chains to one another, giving rise to a complex network (Figure 3.7); it is this cross-linking that gives the wall its mechanical strength. A number of antimicrobial agents exert their effect by inhibiting cell wall synthesis; β-lactam an-tibiotics such as penicillin inhibit the transpeptidases, thereby weakening the cell wall, whilst bacitracin prevents transport of peptidoglycan precursors out of the cell. The action of antibiotics will be discussed further. Although all bacteria (with a few exceptions) have a cell wall containing peptidoglycan, there are two distinct struc-tural types. These are known as Gram-positive and Gram-negative. The names derive from the Danish scientist Christian Gram, who, in the 1880s developed a rapid staining technique that could differentiate bacteria as belonging to one of two basic types . Although the usefulness of the Gram stain was recognised for many years, it
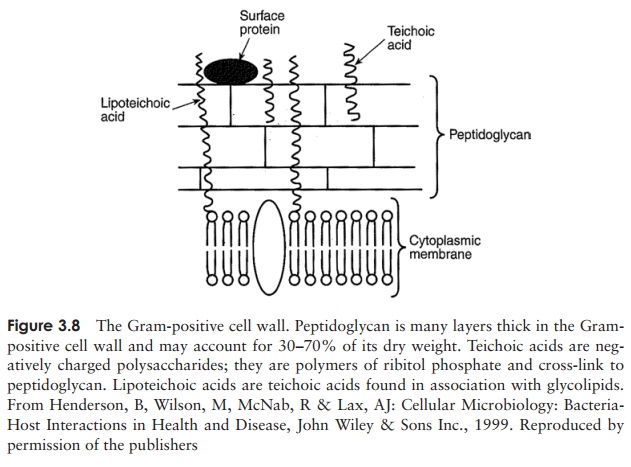
Gram-positive cell walls are relatively simple in structure, comprising several layers of peptidoglycan connected to each other by cross-linkages to form a strong, rigid scaffolding. In addition, they contain acidic polysaccharides called teichoic acids; these contain phosphate groups that impart an overall negative charge to the cell surface. A diagram of the gram-positive cell wall is shown in Figure 3.8.
Gram-negative cells have a much thinner layer of peptidoglycan, making the wall less sturdy, however the structure is made more complex by the presence of a layer of lipoprotein, polysaccharide and phospholipid known as the outer membrane
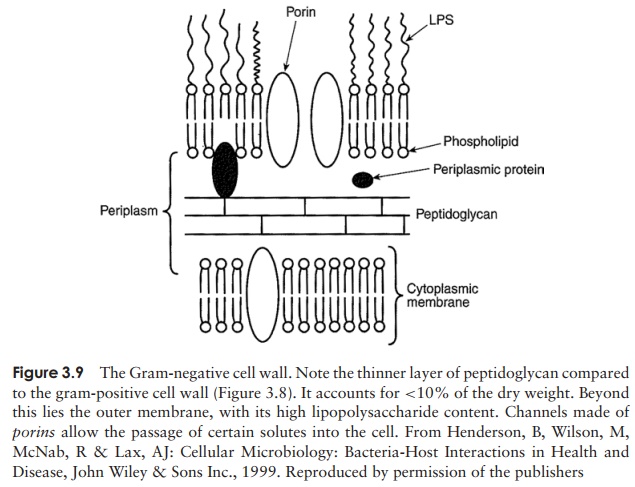
(Figure 3.9). This misleading name derives from the fact that it superficially resembles the bilayer of the plasma membrane; however, instead of two layers of phospholipid, it has only one, the outer layer being made up of lipopolysaccharide. This has three parts: lipid A, core polysaccharide and an O-specific side chain. The lipid A component may act as an endotoxin, which, if released into the bloodstream, can lead to serious con-ditions such as fever and toxic shock. The O-specific antigens are carbohydrate chains whose composition often varies between strains of the same species. Serological meth-ods can distinguish between these, a valuable tool in the investigation, for example, of the origin of an outbreak of an infectious disease. Proteins incorporated into the outer membrane and penetrating its entire thickness form channels that allow the passage of water and small molecules to enter the cell. Unlike the plasma membrane, the outer membrane plays no part in cellular respiration.
Members of the Archaea have a cell wall chemistry quite different to that described above. Instead of being based on peptidoglycan, they have other complex polysaccharides, although a distinction between gram-positive and gram-negative types still occurs.
Beyond the cell wall
A number of structural features are to be found on the outer surface of the cell wall; these are mainly involved either with locomotion of the cell or its attachment to a suitable surface.
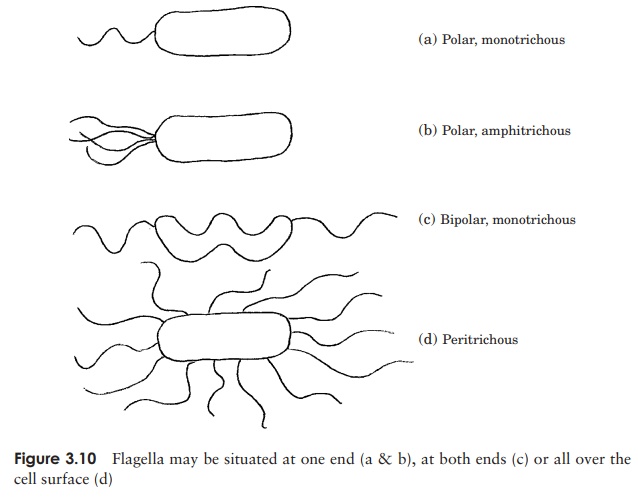
Perhaps the most obvious extracellular structures are flagella (sing: flagellum), thin hair-like structures often much longer than the cell itself, and used for locomotion in many bacteria. There may be a single flagellum, one at each end, or many, depending on the bacteria concerned (Figure 3.10). Each flagellum is a hollow but rigid cylindri-cal filament made of the protein flagellin, attached via a hook to a basal body, which secures it to the cell wall and plasma membrane (Figure 3.11). The basal body com-prises a series of rings, and is more complex in Gram-negative than Gram-positive bacteria. Rotation of the flagellum is an energy-dependent process driven by the basal body, and the direction of rotation determines the nature of the resulting cellular move-ment. Clockwise rotation of a single flagellum results in a directionless ‘tumbling’,
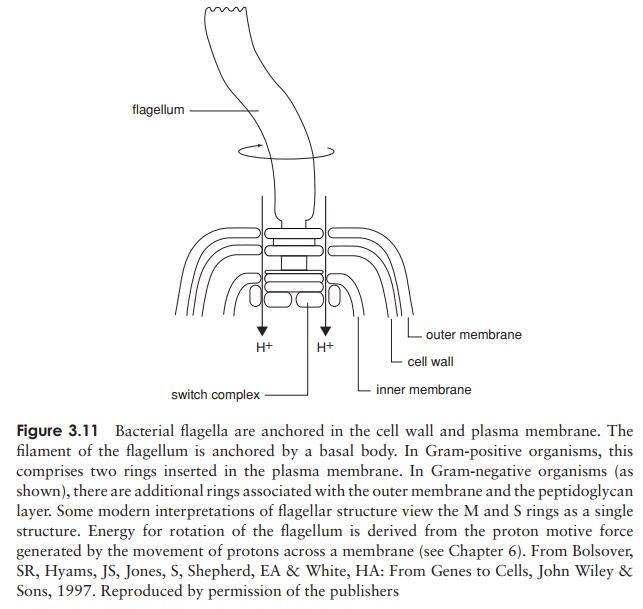
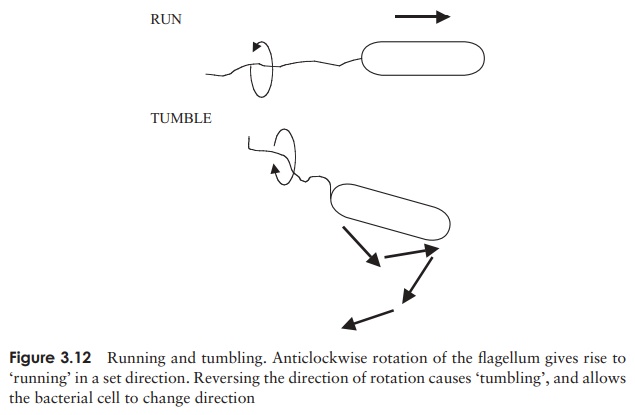
but if it rotates anticlockwise, the bacterium will ‘run’ in a straight line (Figure 3.12). Likewise, anticlockwise rotation causes bunched flagella to ‘run’ by winding around each other and acting as a single structure, whilst spinning in the opposite direction gives rise to multiple independent rotations and tumbling results once more.
Pili (sing: pilus) are structures that superficially resemble short flagella. They differfrom flagella, however, in that they do not penetrate to the plasma membrane, and they are not associated with motility. Their function, rather, is to anchor the bacterium to an appropriate surface. Pathogenic (disease-causing) bacteria have proteins called adhesins on their pili, which adhere to specific receptors on host tissues. Attachment pili are sometimes called fimbriae, to distinguish them from another distinct type of pilus, the sex pilus, which as its name suggests, is involved in the transfer of genetic information by conjugation.
Outside the cell wall, most bacteria have a polysaccharide layer called a glycocalyx. This may be a diffuse and loosely bound slime layer or a better defined, and generally thicker capsule. The slime layer helps protect against desiccation, and is instrumental in the attachment of certain bacteria to a substratum (the bacteria that stick to your teeth are a good example of this). Capsules offer protection to certain pathogenic bacteria against the phagocytic cells of the immune system. Both capsules and slime layers are key components of biofilms, which form at liquid/solid interfaces, and can be highly significant in such varied settings as wastewater treatment systems, indwelling catheters and the inside of your mouth!
Related Topics