Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: A history of fishes
A history of fishes: summary and overview
A history of fishes: summary and overview
As should be obvious, the gaps in our knowledge about fossil fishes and their relationships to one another and to modern groups are large and plentiful. Such gaps are the initiation points for future research. For starters, three topics arise from these unanswered questions and deserve some exploration.
The diversity of fossil fishes
We speak of the success of different ancient groups and compare among them and between modern teleosts and extinct forms. All of the preceding discussion is totally dependent on the fossil record. But how accurately does the fossil record represent the diversity of fossil fishes? How many fishes would we estimate are alive today if we were forced to rely on fossils? As of 1988, approximately 333, or about 8% of modern teleostean genera, are represented by Recent fossils (Carroll 1988; Nelson 1994). Significantly, the number of fossils available for study decreases with time because geological processes tend to destroy fossilized material. Therefore, the fossil record of Recent fishesis at best an optimistic underestimator of the accuracy with which earlier groups are represented.
Fossilization is a chance procedure, compounded by the relatively small surface of the earth available for paleontological discovery. Inadequacy of sampling is obvious when we realize that most fossils are recovered from only the top few meters of rock, and much of the surface land of the Mesozoic and early Cenozoic has been subducted by tectonic
processes. Our limited sample size is aggravated by inaccessibility of major areas of the earth’s surface; recall that 70% of our planet is under water, where very little paleontological exploration occurs. Significantly, about 2400 species, or 10%, of living fishes occur in water deeper than 200 m (Cohen 1970), yet few of the recognized pre-teleostean groups are postulated as having occupied the deep sea. Deepsea fishes, regardless of taxon, are highly convergent in body form and structure; such adaptations should be obvious in fossils and such fishes should be assignable to the deepsea habitat. However, the fossil record for living deepsea groups is understandably limited. For example, stomiiforms are among the most abundant of the deepsea orders, with >50 recognized genera, but only five of these, or about 10%, have a fossil record (Nelson 1984; Carroll 1988) (fossil rarity may also reflect convergence on the trait of reduced ossifi cation, reducing further the likelihood of fossilization). The deep sea is one of the most stable aquatic habitats on earth and it seems unlikely that living in the deep sea is a teleostean innovation. Pre-teleostean diversity in deepsea habitats is obviously underestimated.
Compound these problems further with the realization that many fossil species are described based on a single, often fragmentary, specimen. How many of these fragments remain undiscovered and, more importantly, how many rare species never fossilized? In our search for antecedents of modern groups, how does this selective preservation of forms affect our interpretations of lines of descent, particularly if there exists at best a one-in-10 chance that an ancestor will fossilize? Our optimistic hope is that the fossil record is somehow a proportional and representative subsample of reality, that we accept that we have grossly underestimated the diversity of primitive fishes, and that many more future researchers will take up the challenges of paleontology.
The tangled web of early vertebrate relationships: primitive does not necessarily denote ancestral
It is intellectually frustrating to have major living taxa, e.g. modern agnathans, jawed fishes, and gnathostomes in general, for which we can find no clear ancestral lineages. Such phylogenetic problems beg for solution. As a result, considerable effort has been extended attempting to link modern agnathan groups with Paleozoic forebears, and for that matter, modern gnathostomes with ancestors among the diversity of fishes that proliferated during the Devonian.
Plausible alternative explanations exist. First, the ancestors of modern groups may have died out without leaving fossil remains, at least none that we have found so far. Second, Paleozoic lineages were extinguished, period. Hence similarities between ancient and extant groups result from convergence and perhaps some retention of primitive characteristics derived from a common, distant (unfossilized) ancestor. The latter scenario is perfectly reasonable given the rather advanced condition of bones in the agnathous fishes and of jaws and other supporting bony elements in the early gnathostomes when they first appear in the fossil record. Groups ancestral to these early lineages must have existed for millions of years but lacked the necessary mineralized structures to fossilize. Extinction is a universal characteristic of species; it has been estimated that the average “life span” of a species is around 10 million years (Raup 1988). The mass extinctions that have occurred during the history of life (e.g., the Burgess Shale fauna in the Precambrian, and the Permo-Triassic and Cretaceous– Tertiary extinction events) have been particularly disastrous for shallow marine faunas, wiping out 50–100% of the species in existence at the time (Raup 1988). It seems reasonable to assume that fish lineages were as susceptible to mass extinctions as were contemporaneous invertebrate groups; declines in diversity of actinistians, amiiforms, hybodonts, holocephalans, and perhaps neopterygians at the end of the Cretaceous may attest to the vulnerability of fish groups to mass extinction.
Continuity in fish evolution
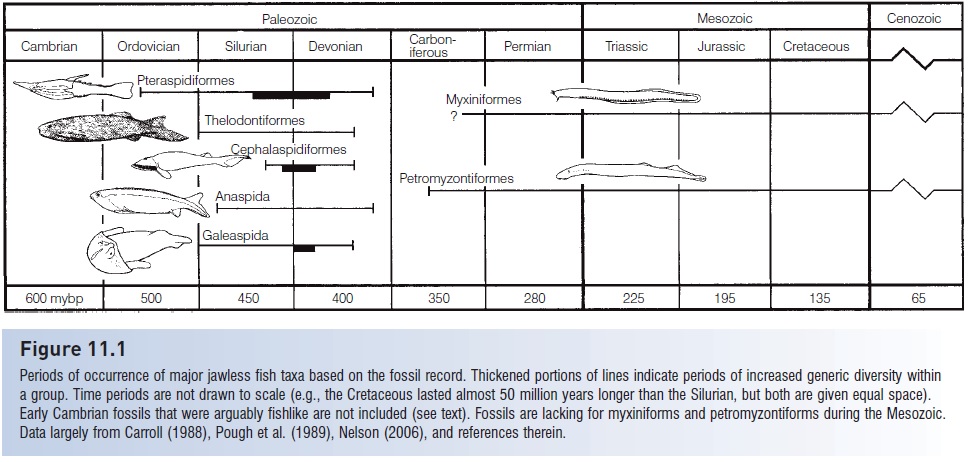
This separation is, however, arbitrary and superfi cial. It is more of a stylistic convenience for organizing a textbook than a statement of philosophy. Students of fish evolution should quickly recognize that modern fishes are extensions of fossil groups. As was pointed out earlier, the majority of modern fish families already existed in the Mesozoic if not earlier (see Fig. 11.1). Although some primitive groups that are unrepresented today (e.g., “ostracoderms”, placoderms, acanthodians, osteolepidiforms, palaeoniscoids) probably deserve separate treatment from modern forms, it makes just as much sense to treat truly ancestral forms, such as primitive dipnoans, actinistians, neopterygians, and chondrichthyans, together with their modern derivatives. To paraphrase paleontologist A. R. McCune, why should mode of preservation – in rocks or in alcohol – be the primary determinant of how we deal with a taxo nomic group? If modern, “primitive” species (e.g., the living coelacanths) were to become extinct through human neglect, would they immediately have to be placed only in a discussion of extinct fishes? It is our hope that students of ichthyology will recognize the continuity that exists between primitive and advanced groups and not view them as separate entities but rather as a continuum of organic change within lineages.
Related Topics