Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: A history of fishes
Subclass Elasmobranchii - Advanced jawed fishes: Chondrichthyes
Subclass Elasmobranchii
Definitive sharklike fossils first appear in the Early Devonian, involving teeth (418 mybp) and an intact shark fossil (409 mybp); scales or dermal denticles are known from the Late Ordovician (455 mybp). The elasmobranchs (“plate or strap gills”) have undergone several major radiations, with much controversy surrounding interrelationships. At least eight orders of elasmobranchs with origins in the Paleozoic arose and disappeared by the Triassic (Compagno 1990b). Identification of the various lineages is based largely on tooth, scale, and spine morphology, and the fossil evidence indicates that, as with bony fishes, foraging and locomotor improvements characterize successive groups.
Elasmobranchs are divided into three infraclasses, the extinct cladoselachimorphs and xenacanthimorphs, and the euselachians, which include modern forms and their extinct relatives. Cladoselachimorphs contained one family, the Cladoselachidae (Fig. 11.26A). Cladoselachids had five gill slits and a terminal mouth. Their dentition, referred to as cladodont, consisted of multicuspid teeth in which the central cusp was usually larger. The teeth were made of enamel-covered dentine and were homologous with scales. These elasmobranchs were often large (2 m), pelagic, marine predators with an unconstricted notochord protected by calcified cartilaginous neural arches, and with small precaudal, lateral keels analogous to those found in modern pelagic sharks (Moy-Thomas & Miles 1971). The dorsal fins were often preceded by a spine that may have been supportive or protective in function. Caudal morphology was functionally symmetrical, although the notochord extended into the dorsal lobe of the fin (Fig. 11.26A). Cladoselachids were recognizably sharklike in appearance.
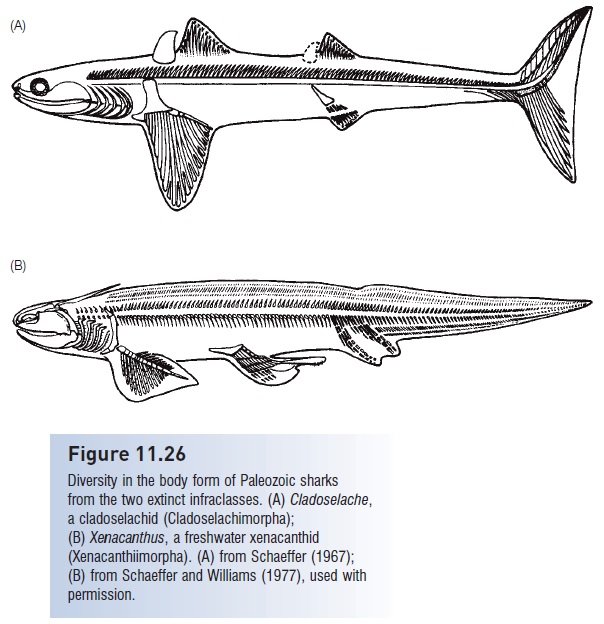
Figure 11.26
Diversity in the body form of Paleozoic sharks from the two extinct infraclasses. (A) Cladoselache, a cladoselachid (Cladoselachimorpha); (B)Xenacanthus, a freshwater xenacanthid (Xenacanthiimorpha). (A) from Schaeffer (1967); (B) from Schaeffer and Williams (1977), used with permission.
Another mainstem elasmobranch infraclass, the xenacanthomorphs, were common in tropical waters from the Lower Devonian into the Triassic. Recognized families include xenacanthids, lebachacanthids, and diplodoselachids. Xenacanths had a tooth type different from the cladoselchids termed pleuracanth, in which the two lateral cusps were large and the median cusp was smaller. Xenacanthids invaded fresh water and assumed an eel-like morphology (Fig. 11.26B). Some xenacanthid sharks had pectoral fins reminiscent of the archipterygium of the dipnoans and may have been bottom dwellers. Xenacanthids also were unusual in possessing two distinct anal fins.
The earliest euselachians were the Ctenacanthiformes of the Middle Devonian to the Upper Triassic. Ctenacanthids had two dorsal fins with prominent spines, an anal fin set far back on the body, and a slightly overhanging snout along with a terminal mouth (Fig. 11.27A). Hybodontiformes of the Triassic and Jurassic are placed in their own division, the Hybodonta, because they are considered the sister group of the modern sharks of the division Neoselachii.
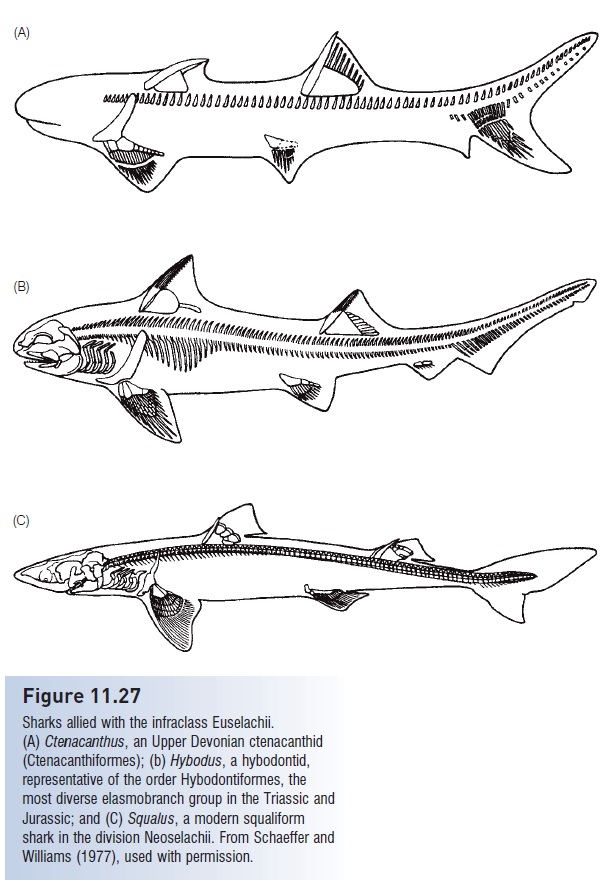
Figure 11.27
Sharks allied with the infraclass Euselachii. (A) Ctenacanthus, an Upper Devonian ctenacanthid (Ctenacanthiformes); (b) Hybodus, a hybodontid, representative of the order Hybodontiformes, the most diverse elasmobranch group in the Triassic and Jurassic; and (C) Squalus, a modern squaliform shark in the division Neoselachii. From Schaeffer and Williams (1977), used with permission.
However, unlike modern neoselachians, hybodonts retained the terminal mouth of the ctenacanth sharks, rather than the subterminal mouth evolved by neoselachians in the Jurassic (Fig. 11.27B). Hybodont teeth represented an innovation over more primitive sharks in that hybodonts had multicuspid teeth that were often differentiated into
Although hybodonts were notably diverse during the Triassic and Jurassic, occupying perhaps as many adaptive zones as modern sharks, neither they nor any of the earlier shark groups survived beyond the Mesozoic. They were replaced by neoselechian modern sharks in marine habitats and by neopterygians in freshwater regions. Neoselachians first appear in the Lower Triassic, contemporaneous with the hybodonts. By the Early Jurassic, recognizably modern sharks are found (Fig. 11.27C). One major distinction between modern and earlier sharks is the characteristic overhanging snout of neoselachians, producing a ventral rather than terminal mouth. The overhanging snout results from an enlarged rostral area that encases a larger olfactory system. Modifications in jaw suspension, jaw–pectoral girdle linkage, and jaw-opening muscles create a protrusible upper jaw and the generation of suction forces, paralleling the trend seen in teleosts. Calcified vertebral centra largely replaced the unconstricted notochord of earlier groups, and fin supports changed from multiple basal cartilages with cartilage radiating out to the fin margins to smaller, fused basal supports (usually three) and fl exible, horny rays termed ceratotrichia supporting the web of the fin. This combination of vertebral and fin modifications should have provided for faster swimming and greater maneuverability.
Regardless of the radiation in question, several elasmobranch innovations probably gave them a selective advantage over the other early gnathostomes present at the time. In contrast to the placoderms and most acanthodians, sharks quickly evolved a tooth replacement mechanism. Teeth grew in whorls or spiral bands (Fig. 11.28), with the functional, exposed tooth backed up by several replacement teeth embedded in the jaw cartilage. As embedded teeth grew, they moved along the whorl until they erupted at the jaw periphery, only to be later replaced by younger teeth. Dentition replacement patterns differ among different lineages of modern sharks, but in all

Figure 11.28
Tooth replacement in chondrichthyans. (A) Cross-section through the jaw of a modern shark, showing a functional tooth backed by rows of developing replacement teeth. Variations on this mechanism are found in many fossil groups. (B) Symphysial (middle) portion of the lower jaw of the late Paleozoic edestoid Helicoprion, thought to be a holocephalan, showing its spiral replacement tooth whorl. After Carroll (1988) and Pough et al. (1989).
Other characteristics of modern sharks undoubtedly had their origins in late Paleozoic groups. Sexually dimorphic males had pelvic fins modified as intromittent organs for sperm deposition, indicating that internal fertilization evolved early in chondrichthyans (see Fig. 11.26B). A strong dependence on electroreception, highly acute olfactory capabilities (associated with the longer snout that houses the nasal capsules), increased buoyancy through oil accumulation in the liver (paralleling gas bladder evolution in bony fishes), and large brain and body size have all contributed to the position of modern elasmobranchs among the top predators in marine habitats.
Related Topics