Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: A history of fishes
Class Actinopterygii - Advanced jawed fishes : teleostomes (Osteichthyes)
Class Actinopterygii
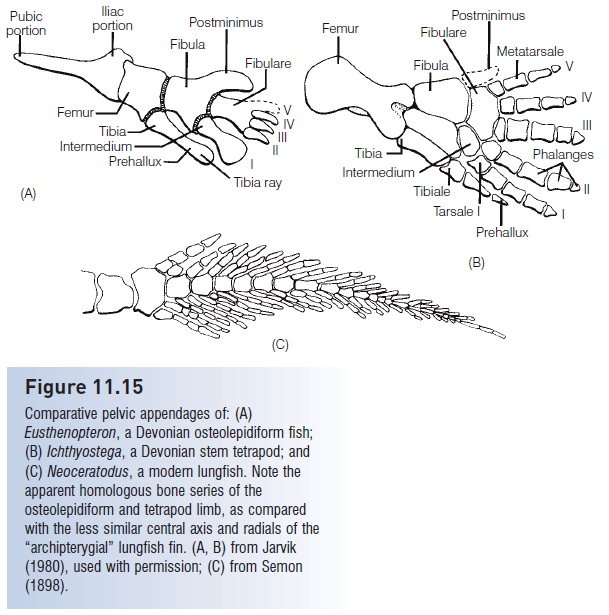
The primitive fish groups discussed so far are interesting for their antiquity and diversity, and for the effort required by paleontologists to slowly unearth and interpret features of their design. Yet these fishes bear little resemblance to most modern groups and are at most only distantly related to the familiar fishes of today. Speculation about the natural history, behavior, and ecology of extinct forms is based on scant information, much of it difficult to interpret. It is consequently challenging to “imagine” these animals as the living creatures that they were. These difficulties do not apply however to the ancestors of the Actinopterygii, the most successful of today’s fishes. Although just as ancient as most of the other groups, primitive ray-finned fishes are similar in size and shape to many extant fishes, and many of their fossils are very well preserved. We can therefore equate many fossil and extant actinopterygians in terms of descendancy, form, and possibly function.
Subclass Cladistia
After a great deal of effort and no small amount of controversy (see Nelson 1994; Helfman et al. 1997), the weight of opinion has shifted to recognize the modernpolypteriforms as a separate subclass and the sister group of the other two subclasses of actinopterygians, the Chondrostei and Neopterygii. Also referred to asbrachiopterygians, fossil cladistians are known only as far back as the Middle Cretaceous of Africa and Late Cretaceous of South America. This represents a dramatic gap in the fossil record for a group considered more primitive than other actinopterygians, which are known from the Devonian (cheirolepidiform and palaeonisciform chondrosteans) and the Triassic (semionotiform
Subclass Chondrostei
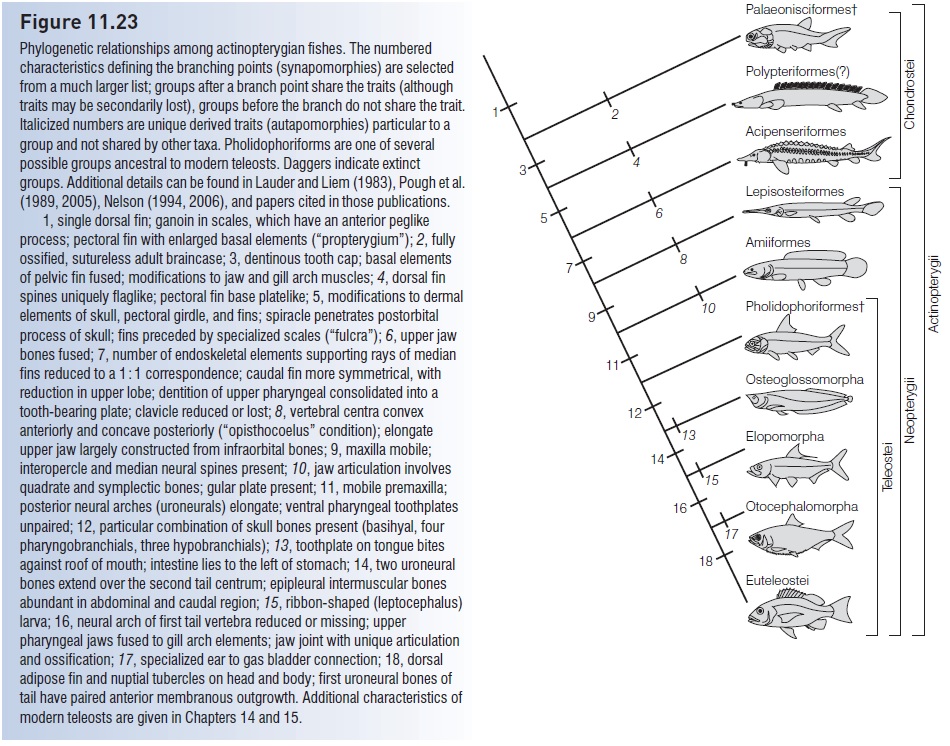
The origins of the Actinopterygii are once again obscure. Scale fragments appear in Late Silurian marine deposits, which may mean that the group is older than the sarcopterygians and as old as placoderms and elasmobranchs. Only the acanthodians among the bony, jawed fishes are of greater antiquity, supporting speculation of an acanthodian ancestry for modern bony fishes. However, complete fossil actinopterygians do not appear until the Mid to Late Devonian, when the group had expanded into a variety of marine, estuarine, and freshwater habitats. These early fishes, collectively known as palaeoniscoids (but see below) were relatively small (5–25 cm) and were distinguished from sarcopterygians by the presence of a single triangular dorsal fin, a forked heterocercal tail with no upper lobe above the unconstricted notochord, paired fins with narrow rather than fleshy bases, dermal bones lacking a cosmine layer, scales joined by a peg-and-socket arrangement and covered with ganoine (“ganoid” scales), relatively large eyes, and a blunt head (Fig. 11.19A, B). The term “ray-fin” refers to the parallel endoskeletal fin rays that were derived from scales. These rays supported the median and paired fins, which were moved by adjacent body musculature. In contrast, the fins of the Sarcopterygii had a thick, bony central axis and muscles contained in the fin itself (see Fig. 11.15C). Chondrosteans include one extant and 10 extinct orders, with relationships obscure. The most primitive group, the Devonian cheirolepidiforms, includes a species with the distinction of possessing the largest number of pelvic fin rays known among fishes, living or otherwise. Cheirolepis canadensis had 124 such rays versus six or fewer in living teleosts. The most diverse order, the Palaeonisciformes, contained four suborders and 17 families of wellrepresented fishes that showed tremendous morphological diversity (Figs 11.19, 11.20). The other orders of early chondrosteans are often lumped together as “palaeoniscoids” despite taxonomic differences, and palaeoniscoids are then treated as ancestral to later neopterygians and therefore teleosts.
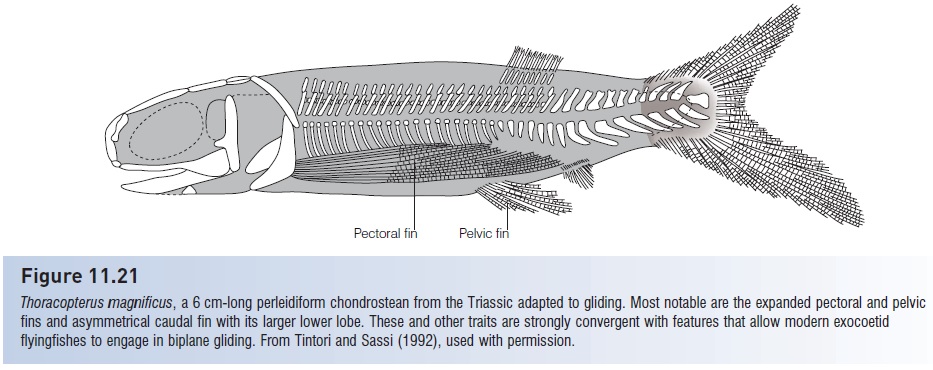
Among the other orders are the Carboniferous tarrasiiforms, which were remarkably convergent with many modern eel-like forms, possessing an elongate body, dorsal and anal fins continuous with the caudal fin (the latter being diphycercal in this group), and pelvic fins and scales reduced or absent (Fig. 11.20). Saurichthyiforms converged on a needlefish body shape and are thought to have been similarlypredatory on small fishes, and phanerorhynchiforms bore a superficial resemblance to modern sturgeons. An advanced order from the Late Triassic, the perleidiforms, included Thoracopterus, a genus with expanded paired fins thought capable of biplane gliding, as occurs in modern exocoetid flyingfishes (Tintori & Sassi 1992). Thoracopterus possessed the enlarged pectoral and pelvic fins, reinforced rays in the paired fins, asymmetrical caudal fin, expanded caudal neural spines for muscle insertion, posterior position of dorsal and anal fins, and head shape of
Such convergence, remarkable in itself, would have required substantial reduction in the otherwise heavy armoring characteristic of the early chondrosteans. Modern chondrosteans, the acipenseriform sturgeons and paddlefishes, have fossil representation in the Jurassic and Lower Cretaceous, respectively.

Figure 11.16
Dorsal (A) and lateral (B) views of the reconstructed elpistostegalian fish, Tiktaalik roseae. Features evident are the lack of opercular bones, the tetrapod-like arrangement of elements in the pectoral fins/limbs, and the stout ribs forming a rib cage that may have protected the lungs. From Daeschler et al. (2006), used with permission.
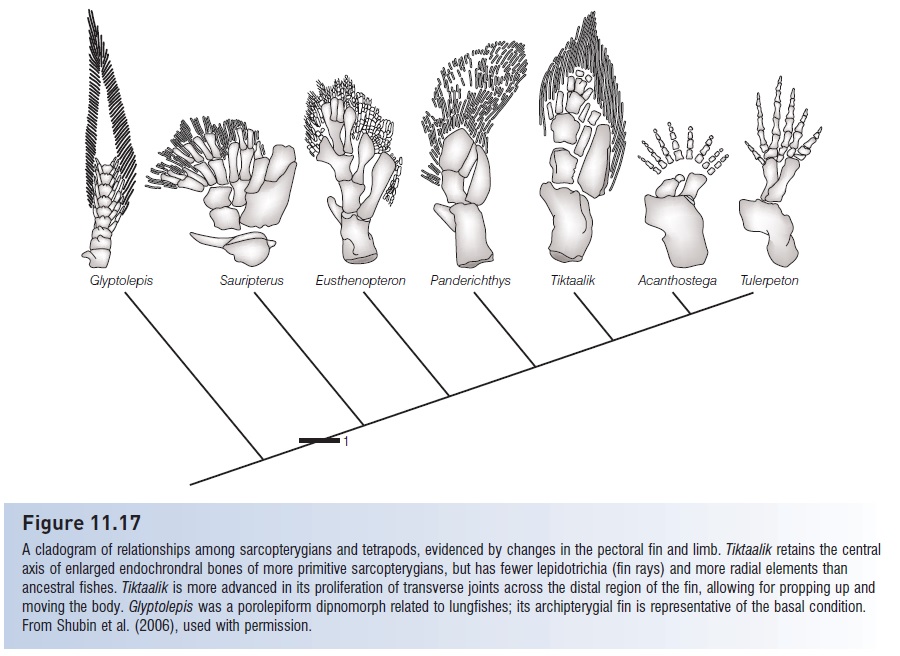
Figure 11.17
A cladogram of relationships among sarcopterygians and tetrapods, evidenced by changes in the pectoral fin and limb. Tiktaalik retains the central axis of enlarged endochrondral bones of more primitive sarcopterygians, but has fewer lepidotrichia (fin rays) and more radial elements than ancestral fishes. Tiktaalik is more advanced in its proliferation of transverse joints across the distal region of the fin, allowing for propping up and moving the body. Glyptolepis was a porolepiform dipnomorph related to lungfishes; its archipterygial fin is representative of the basal condition. From Shubin et al. (2006), used with permission.
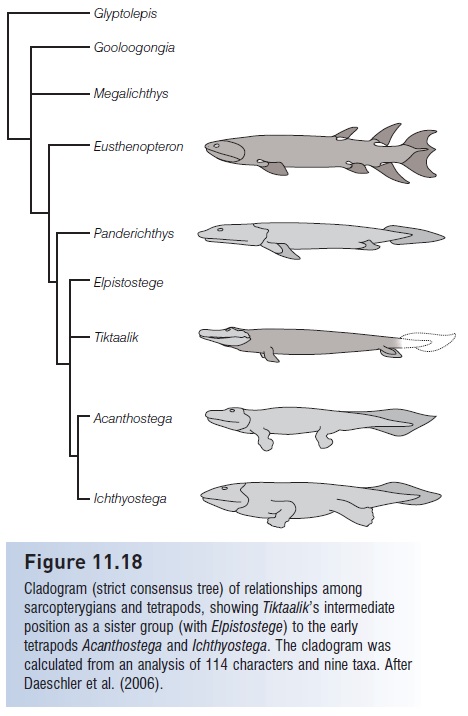
Figure 11.18
Cladogram (strict consensus tree) of relationships among sarcopterygians and tetrapods, showing Tiktaalik’s intermediate position as a sister group (with Elpistostege) to the early tetrapodsAcanthostega and Ichthyostega. The cladogram was calculated from an analysis of 114 characters and nine taxa. After Daeschler et al. (2006).

Figure 11.19
Actinopterygian fishes at different grades of development. (A) Moythomasia and (B) Mimia, two primitive palaeoniscoid fishes from the Upper Devonian, with thick rhomboidal scales extending onto the fins, broadly triangular dorsal and anal fins, fulcral (ridge) scales along the back, a long mouth, and an asymmetrical heterocercal tail. (C) Parasemionotus, a pre-teleostean neopterygian from the Triassic, showing more flexible fins, shorter mouth, and abbreviate heterocercal tail. (D) Eolates, an advanced euteleost from the Lower Eocene, with characteristic teleostean diversified dorsal and anal fins, shortened vertebral column, premaxillary dominated upper jaw, and homocercal tail. (A) after Jessen (1966); (B) after Gardiner (1984); (C) after Lehman (1966); (D) after Sorbini (1975).

Figure 11.20
Morphological (and ecological) convergence in fish evolution. Palaeoniscoids were ancestral to early neopterygians, which were ancestral to modern teleosts. Certain body designs or plans have apparently been repeatedly favored in actinopterygians, leading to convergent designs among unrelated lineages. These striking convergences in body shape and presumably function are depicted for representative palaeoniscoids, early neopterygians, and teleosts. 1, Elongate piscivores with long, ooth-studded jaws and dorsal and anal fins placed posteriorly for rapid starts; 2, compressed-bodied, predatory, shallow water fishes with deeply forked tails and trailing fins; 3, broad-finned bottom feeders with subterminal mouths; 4, eel-like benthic forms; and 5, compressed, circular forms with large fins for maneuverability in shallow water habitats with abundant structure. Gliding fishes such as the Triassic chondrostean Thoracopterus (Fig. 11.21) can also be equated with modern teleostean flyingfishes. Adapted from Pough et al. (1989), not drawn to scale.
Related Topics