Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: A history of fishes
Jawless fishes - A history of fishes
Jawless fishes
The very first fishlike vertebrates undoubtedly evolved from invertebrates, perhaps a cephalochordate. However, the first “fishes” left no fossil record and their form and relationships remain a mystery. By the time fishlike fossils appear in Early Cambrian deposits, roughly 530 million years before present (mybp) (Fig. 11.1), complex tissue types had evolved, including filamentous gills, V-shaped myomeres, and a distinct dorsal fin.
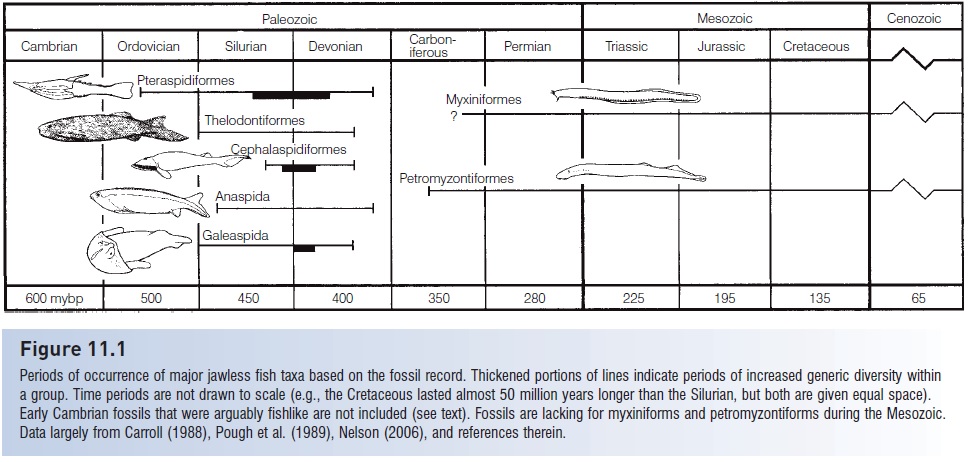
Figure 11.1
Periods of occurrence of major jawless fish taxa based on the fossil record. Thickened portions of lines indicate periods of increased generic diversity within a group. Time periods are not drawn to scale (e.g., the Cretaceous lasted almost 50 million years longer than the Silurian, but both are given equal space). Early Cambrian fossils that were arguably fishlike are not included (see text). Fossils are lacking for myxiniforms and petromyzontiforms during the Mesozoic. Data largely from Carroll (1988), Pough et al. (1989), Nelson (2006), and references therein.
The very first fishlike vertebrates undoubtedly evolved from invertebrates, perhaps a cephalochordate. However, the first “fishes” left no fossil record and their form and relationships remain a mystery. By the time fishlike fossils appear in Early Cambrian deposits, roughly 530 million years before present (mybp) (Fig. 11.1), complex tissue types had evolved, including filamentous gills, V-shaped myomeres, and a distinct dorsal fin. New dis- coveries are made almost annually, but the currently recognized oldest species,Myllokunmingia fengjiaoa, was found in the Chengjiang geological formation of Yunnan
Province in southwestern China. Myllokunmingia was 3– 4 cm long and is thought to be allied with (is a sister group to) ancestors of modern lampreys, although agreement is far from universal (Xian-guang et al. 2002; Shu et al. 2003).
If modern cephalochordates such as the lancelets (Branchiostoma) are considered fishlike – if not exactly fishes – then the ancestry of fishes can be traced farther back to the cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon) from the Lower Cambrian, or to the muchheralded Pikaia with its dorsal nerve cord and notochord, from the Middle Cambrian Burgess Shale of British Columbia (see Amphioxiforms).
Clearly recognizable fish specimens, such as the arandaspid pteraspidomorph Sacabambaspis janvieri from Bolivia, appear later, dating to 470 mybp (Gagnier et al. 1986; Gagnier 1989). This and related jawless, finless forms inhabited shallow seas or estuarine habitats in tropical and subtropical regions of the Gondwanan and Laurasian supercontinents. Their innovations include true bone(probably evolved independently in several ancestral groups) and a muscular feeding pump. The former adaptation, which existed only as an external covering, would have provided protection from predators to which softer ancestors were more vulnerable, as well as serving as a metabolic reserve for calcium and phosphate and an insulator of electrosensory organs (Northcutt & Gans 1983; Carroll 1988). A muscular feeding pump would have been more efficient for moving food-bearing water through a filtration mechanism than was the ciliary feeding mechanism of protochordates. Another major advance over the cephalochordates that preceded them was that, although lacking jaws, the early fossilized fishes were craniates. They had a head region containing a brain with specialized sensory capsules and cranial nerves, all contained in a protective skeletal braincase (Maisey 1996).
Related Topics