Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: A history of fishes
Class Sarcopterygii - Advanced jawed fishes I: teleostomes (Osteichthyes)
Class Sarcopterygii
Ancestral sarcopterygians remain one of the most actively studied fossil groups of fishes, in no small part because of their place in tetrapod evolution. In recent years, abundant discoveries have been made, often prompting reanalysis of relationships among fossil and extant groups. Agreement is far from universal: Forey (1998) summarized major hypotheses, presenting 13 different phylogenies proposed by different authorities in the last 20 years of the 20th century. Forey’s concluding analysis is presented in Fig. 11.10 and is largely followed here. The debate revolves largely around
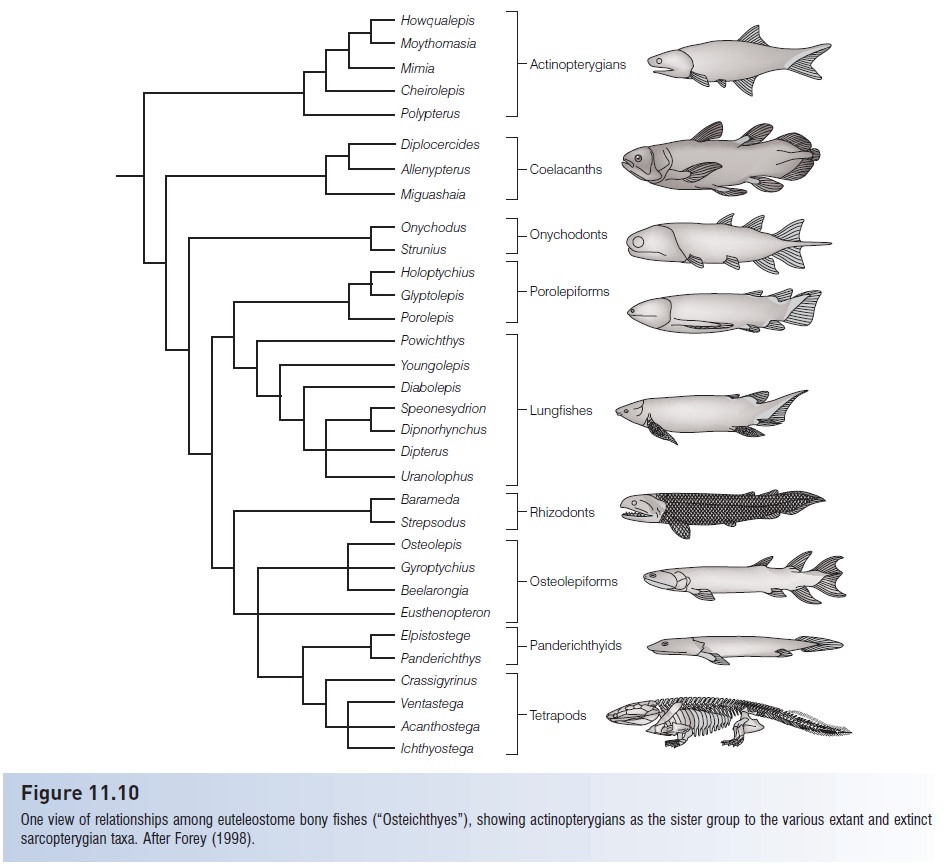
Figure 11.10
One view of relationships among euteleostome bony fishes (“Osteichthyes”), showing actinopterygians as the sister group to the various extant and extinct sarcopterygian taxa. After Forey (1998).
Until the dust settles, many higher taxonomic groupings omitted from this treatment are listed as “unranked” and are given numerical designations 1a–4b in Nelson (2006). The exception is Tetrapodomorpha, which we include as “unranked” between class and infraclass. Most of the omitted taxa fall between the offi cial rankings of subclass and superorder, but a formal designation does not yet exist. We fully expect that in the next few years, researchers in this exciting area will erect names and ranks for these groups, alleviating the confusion that currently plagues a student first encountering the admittedly bewildering array of unfamiliar names and serial numbers.
Subclass Coelacanthimorpha, order Coelacanthiformes
Fossil coelacanthimorphs (or Actinistia) appeared in the Middle Devonian and are not known after the Late Cretaceous. They occurred worldwide in both marine and fresh water. The fossil record of the group is extensive: at least 83 valid species in 24 genera and perhaps nine families are recognized. Diversity was maximal during the Early Triassic, when 16 described species existed (Forey 1998) (Fig. 11.11). All but one family and two species are extinct.
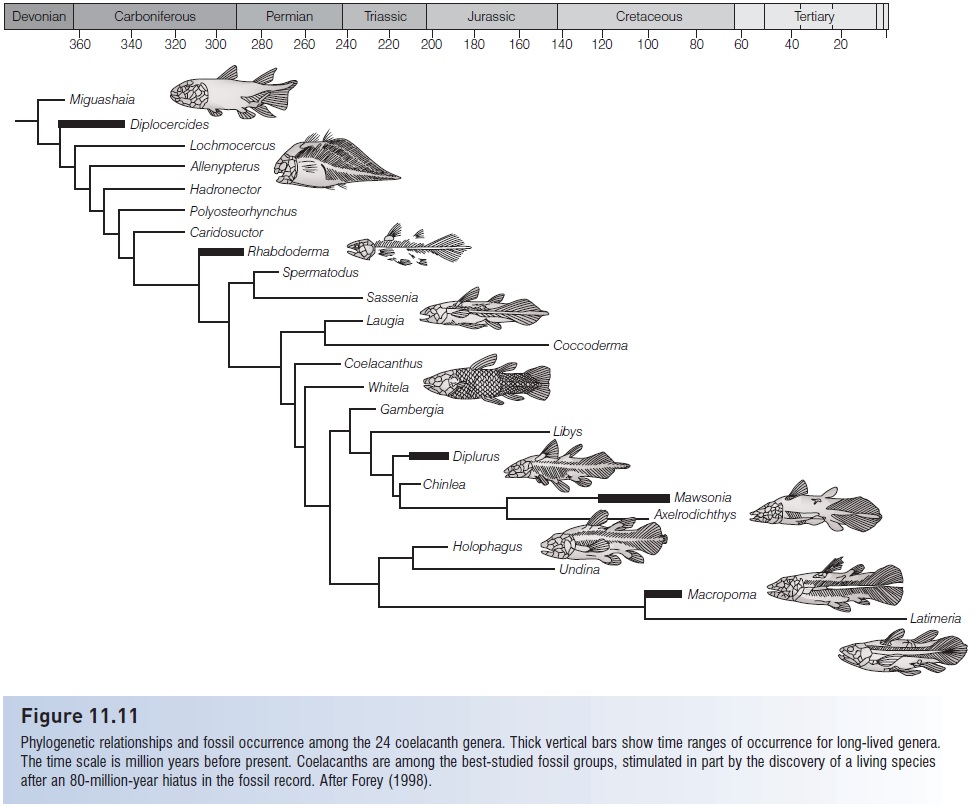
Figure 11.11
Phylogenetic relationships and fossil occurrence among the 24 coelacanth genera. Thick vertical bars show time ranges of occurrence for long-lived genera. The time scale is million years before present. Coelacanths are among the best-studied fossil groups, stimulated in part by the discovery of a living species after an 80-million-year hiatus in the fossil record. After Forey (1998).
Coelacanths are in many respects more specialized than other sarcopterygians, possessing a unique spiny rather than a lobate first dorsal fin; a three-lobed caudal fin with a middle fleshy, fringed lobe (the term “coelacanth” describes the hollow nature of the fin rays that support the tail); a rostral organ involving a rostral cavity with several openings on the snout associated with electroreception; and lacking internal choanae, cosmine in the scales, branchiostegals, and a maxilla. Most evolution in the group occurred during the Devonian (some early genera had a heterocercal rather than a diphycercal tail), and later species are surprisingly unchanged in body shape and jaw morphology from the early representatives, although trends of change (reduction in some bones, increases in others) have occurred (Cloutier 1991). Rapid evolution and morphological variation occurred in early coelacanths, including an eel-like species from the Middle Devonian (Friedman & Coates 2006). Prior to the discovery of the living Coelacanth in 1938 (see The living coelacanths, at least for now), coelacanthimorphs were of interest primarily to paleontologists as a specialized, extinct group notable for its conservativism and its relationship to the reputed ancestors of tetrapods.
Subclass Dipnoi (Dipnotetrapodomorpha)
Besides coelacanths, sarcopterygians consist of two other subclasses: (i) the dipnoans (also called dipnotetrapodomorphs), consisting of a variety of extinct fishes with stout bodies and paddlelike paired fins (including the specialized, modern lungfishes); and (ii) the Tetrapoda that emerged onto land to become amphibians, reptiles, birds, and mammals. Major groups within the Dipnoi are the dipnomorphs (two fossil superorders aligned to lungfishes and the lungfish superorder itself) and the tetrapodomorphs (including the rhizodontomorphs, osteolepidomorphs, and the infraclass Elpistostegalia).
Dipnomorphs
Dipnomorphs are an unranked taxon made up of extinct fishes in the superorder Porolepimorpha (one order, two families from the Devonian), plus two superorders of lungfishes, the extinct Dipterimorpha (10 families) and extant Ceratodontimorpha (one order, three extant families, withsome extinct genera). Lungfishes as a group have been generally referred to as Dipnoi or dipnoans (“doublebreathing”). Dipteramorphs arose in the Devonian in marine environments, expanded into freshwater habitats, and died out by the end of the Triassic. Primitive lungfishes were characterized by: two dorsal fins; fleshy, scale-covered, paired, leafl ike archipterygial fins with a bony central axis and with fin rays coming off the central axis; a lack of teeth on the marginal jaw bones, but with tooth plates inside the mouth, and with the premaxilla, maxilla, and dentary missing; a solid braincase; and a pore-filled, cosmine coating on the dermal bones that covered the skull and scales and that may have been associated with electroreception (Fig. 11.12). Later species occupied fresh water, and trends in lungfish evolution include loss of the first dorsal fin, fusion of the median fins (second dorsal, caudal, anal) to form a symmetrical tail (earlier forms had heterocercal tails), elaboration of the tooth plates and development of replaceable dentition, replacement of ossified centra with cartilage, fusion of skull bones, and concomitant loss of the cosmine covering.

Figure 11.12
Extinct and extant lungfishes. (A) Scaumenacia, an Upper Devonian lungfish from eastern Canada; (B) Toothplates from a fossil lungfish, Ceratodus, from the Upper Triassic (c. 5 cm wide) and from the extant Australian lungfish, Neoceratodus (upper structure). The Australian lungfish is considered to be more similar to ancestral forms than are the living African and South American species. TheNeoceratodus toothplate is mounted on a piece of modeling clay. (A) from Jarvik (1980), used with permission; (B) photo by G. Helfman.
Ceratodontimorphs appear first in the Lower Triassic and are represented today by the freshwater order Ceratodontiformes, containing three families and six species of lungfishes in Australia (Ceratodontidae, one species), South America (Lepidosirenidae, one species), and Africa (Protopteridae, four species). Modern lungfishes take the anatomical trends to the extreme, having eel-like, largely cartilaginous bodies, lacking any cosmine bony layers, and possessing diphycercal tails. The modern Australian lungfish is more similar to the heavier bodied dipnoans of the Paleozoic and Mesozoic. Although limited to fresh waters on three continents today, fossil ceratodontids occupied North and South America, Africa, and Madagascar, many in marine deposits.
Lungfishes underwent extensive diversifi cation during the Devonian, evolving more than 60 genera and 100 species, 80% of which occurred during the Upper Devonian (Marshall 1987). Numbers diminished substantially during the Carboniferous. Many lungfish species are known only from fossilized toothplates, with toothplates and other structures found in fossilized lungfish burrows. These finds indicate that air breathing and estivation (entering torpor and burrowing in mud during drought) evolved as early as the Devonian, a fortuitous (for paleontologists) instance of fish waiting for rains that never came (Moy-Thomas & Miles 1971). Some ceratodontids were quite large; a North American Jurassic species, Ceratodus robustus, was 4 m long and may have weighed as much as 650 kg (Robbins 1991). The modern genus Neoceratodusoccurs as early as the Upper Cretaceous in Australia. The lepidosirenid lungfishes of Africa and South America represent a family that goes back to the Late Carboniferous, but members of the two extant genera do not appear until the Eocene and Miocene, on the same continents where they occur today (Carroll 1988).
Much controversy has swirled around the ancestry of lungfishes, as well as a possible dipnoan ancestry for terrestrial vertebrates (see reviews in Carroll 1988; Pough et al. 1989). Some of this speculation originated with the early misidentification of lungfishes as amphibians. More recent arguments have focused on shared aspects of the lungs, limblike fins, and internal nostrils (e.g., Rosen et al. 1981). However, workers in this area have increasingly reached the conclusion that the ancestry of tetrapods is more closely linked to another group of sarcopterygians, the infraclass Elpistostegalia.
Related Topics