Chapter: Biochemical Pharmacology : Drugs that act on sodium and potassium channels
Structure and function of synapses
Structure and function of
synapses
As we have seen, the
presynaptic action potential will open voltage-gated calcium channels and
thereby trigger exocytosis of the neurotransmitter that is stored in vesicles.
The transmitter will then bind to a postsynaptic receptor. This will typically
result in a local change to the postsynaptic membrane potential, which may or
may not trigger a com-plete action potential (Figure 7.3). While a great many
dif-ferent transmitters exist, individual neurons only seem to be using very
few different ones. Although the time-honoured textbook dogma of one
transmitter per neuron only is no longer valid, we will, for the purpose of
this class, pretend it to be.
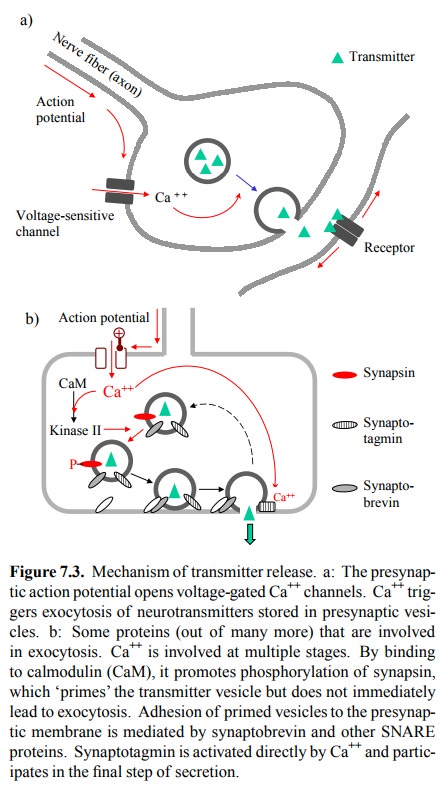
Calcium promotes transmitter
exocytosis at multiple stages. There are several pools of neurotransmitter
vesicles in the nerve ending, which differ by their maturity (e.g., amount of
transmitter stored) and their availability for immediate exocytosis. One effect
of calcium consists in the recruitment of vesicles from an immobile, cell
skeleton-attached pool into the mobile pool. This is achieved by
calmodulin-dependent phosphorylation of synapsin, a protein associat-ed with
the cytosolic surface of the vesicle membrane. Mo-bilized vesicles will attach
themselves to the cytoplasmic membrane. Attachment is mediated by mutual
recognition of so-called SNARE membrane proteins located on both the vesicle
and the cytoplasmic membrane. One such pro tein - synaptobrevin - is the target
of tetanus and botulinum toxins.
The very fast release of
neurotransmitter (1 millisecond after presynaptic excitation) is due to the
exocytosis from pre-attached vesicles. This is effected via synaptotagmin,
which binds and directly responds to calcium. Besides the proteins mentioned,
there is a somewhat distressing multi-tude of additional proteins also
participating in neurotrans mitter release. The precise roles of most of these
proteins remain unsettled.
Related Topics