Chapter: Biochemical Pharmacology : Drugs that act on sodium and potassium channels
Potassium channel blockers
Potassium channel blockers
Among the potassium channels,
it is not the voltage-gated ones but a ligand-gated channel that constitutes
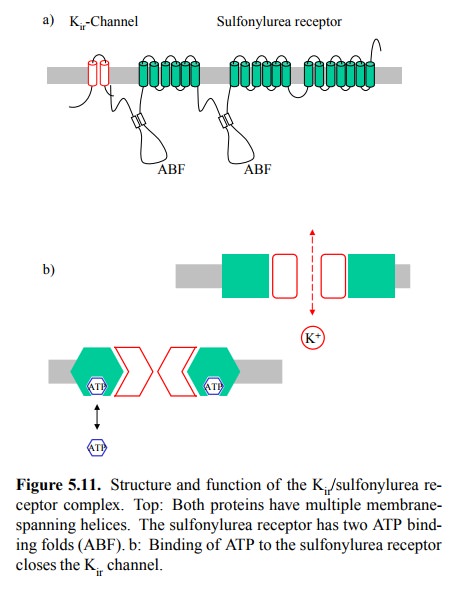
the main drug target. This is the ATP-sensitive `inward rectifier' (Kir)
channel8. The ATP sensitivity is conferred by a second membrane
protein with which it is associated, the so-called sulfonylurea receptor
(Figure 5.11a). The sulfonylurea re-ceptor is homologous to the family of `ABC'
(ATP-binding cassette)
transporters that occurs in both prokaryotes and eukaryotes. These proteins mediate the ATP-driven mem-brane
transport of a wide variety of substances, including the extrusion of toxic
compounds9. The sulfonylurea recep-tor, however, serves a different
purpose: The conforma-tional change induced by binding of ATP is relayed to the
K+ channel, which thus becomes responsive to ATP: High levels of ATP
inhibit the channel, while lower levels cause it to open. This is schematically
depicted in Figure 5.11b.
The sulfonylurea receptor
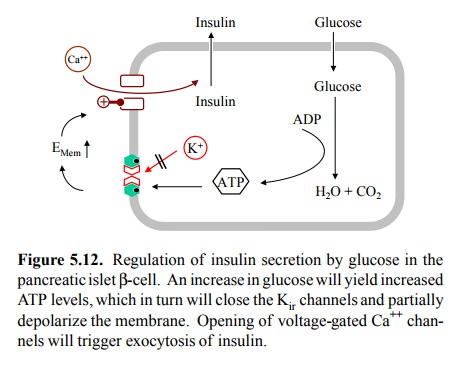
does not only occur in nerve and muscle cells but also in pancreatic β-cells. These are located in the islets of Langerhans, and they
function as both sensors of glucose and as purveyors of insulin. Insulin
secretion is triggered by membrane depolarization, so that these cells are in
fact excitable cells. The control of insulin secretion, and the role of the Kir
channel in it are depicted in Figure 5.12.
The `message' conferred by
insulin is that glucose is plen-tifully available, and that accordingly all
cells should in-crease their uptake, glycogen stores should be replenished, and
internal glucose production (gluconeogenesis) should shut down. Accordingly, a
high level of blood glucose should stimulate insulin secretion. This is
accomplished as follows: High blood glucose will raise the level of glu-cose in
the β-cell. Glucose breakdown will raise the level of ATP, and the
subsequent closure of the Kir channel will induce membrane
depolarization. This, in turn, will open voltage-gated Ca++
channels, and the increase in intracellu-lar Ca++ will trigger
exocytosis of insulin stored in secretory vacuoles. The same mechanism of
coupling between membrane depolarization and exocytosis applies to the release
of neurotransmitter in a synapse in response to a presynaptic action potential
(see later).
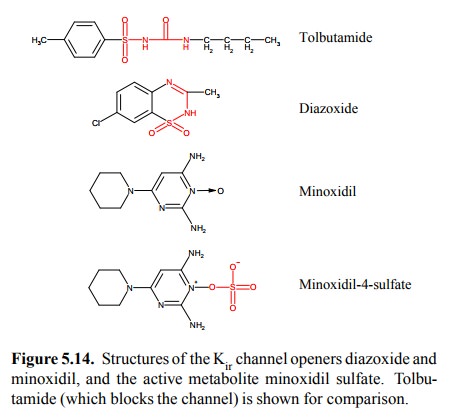
Sulfonylurea derivatives such
as tolbutamide (Figure 5.14) bind to the sulfonylurea receptor and thereby
promote the closing of the Kir channel. This amplifies the secretion
of insulin – a desirable effect in type II diabetics, who suffer not from an
absolute lack of insulin (as type I diabetics do) but from an inadequate
response to it. Sulfonylurea drugs can be used orally, which is more convenient
than insulin therapy. It is also less prone to result in excessive insulin
action (hypoglycemia), since the physiological signal is amplified but not
substituted. However, commonly the β-cells `wear out' eventually, and insulin
therapy has to be instituted anyway.
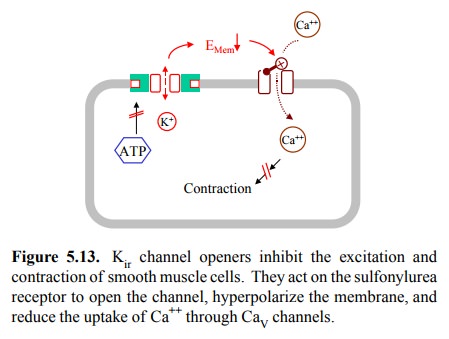
Another, as yet experimental
use of sulfonylurea deriva-tives targets the Kir channels in the
heart in certain forms of arrhythmias. The sulfonylurea receptors in the heart
are somewhat different from those in the β-cells, which makes selective drug action
possible. Closing potassium chan-nels should increase the excitability of heart muscle cells – in contrast to the effect of sodium (and
calcium) channel blockers! Don't worry, though – the cardiologists will find a
way to show us why this is a good thing, like old farm ers have a saying to
predict each kind of weather in ret-rospect.
Related Topics