Sexual Reproduction in Plants - Female reproductive part - Gynoecium | 12th Botany : Chapter 1 : Asexual and Sexual Reproduction in Plants
Chapter: 12th Botany : Chapter 1 : Asexual and Sexual Reproduction in Plants
Female reproductive part - Gynoecium
Pre-fertilization structure and events
The hormonal and
structural changes in plant lead to the differentiation and development of
floral primordium. The structures and events involved in pre-fertilization are
given below
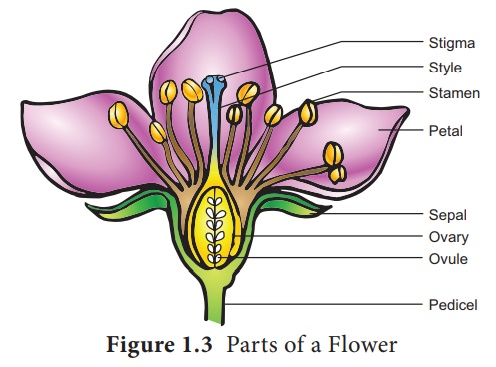
Female reproductive part – Gynoecium
The gynoecium
represents the female reproductive part of the flower. The word gynoecium
represents one or more pistils of a flower. The word pistil refers to the
ovary, style and stigma. A pistil is derived from a carpel. The word ovary
represents the part that contains the ovules. The stigma serves as a landing
platform for pollen grains. The style is an elongated slender part beneath the
stigma. The basal swollen part of the pistil is the ovary. The ovules are
present inside the ovary cavity (locule)on the placenta.
Gynoecium (carpel)
arises as a small papillate outgrowth of meristematic tissue from the growing
tip of the floral primordium. It grows actively and soon gets differentiated
into ovary, style and stigma. The ovules or megasporangia arise from the
placenta. The number of ovules in an ovary may be one (paddy, wheat and mango)
or many (papaya, water melon and orchids).
Structure of ovule(Megasporangium):
Ovule is also called megasporangium and is protected by one or two covering called integuments. A mature ovule consists of a stalk and a body. The stalk or the funiculus (also called funicle) is present at the base and it attaches the ovule to the placenta. The point of attachment of funicle to the body of the ovule is known as hilum. It represents the junction between ovule and funicle. In an inverted ovule, the funicle is adnate to the body of the ovule forming a ridge called raphe. The body of the ovule is made up of a central mass of parenchymatous tissue called nucellus which has large reserve food materials. The nucellus is enveloped by one or two protective coverings called integuments. Integument encloses the nucellus completely except at the top where it is free and forms a pore called micropyle. The ovule with one or two integuments are said to be unitegmic or bitegmic ovules respectively. The basal region of the body of the ovule where the nucellus, the integument and the funicle meet or merge is called as chalaza.
There is a large, oval, sac-like structure in the nucellus toward the micropylar end called embryo sac or female gametophyte. It develops from the functional megaspore formed within the nucellus. In some species(unitegmic tenuinucellate) the inner layer of the integument may become specialized to perform the nutritive function for the embryo sac and is called as endothelium or integumentary tapetum (Example : Asteraceae). There are two types of ovule based on the position of the sporogenous cell. If the sporogenous cell is hypodermal with a single layer of nucellar tissue around it is called tenuinucellate type. Normally tenuinucellate ovules have very small nucellus. Ovules with subhypodermal sporogenous cell is called crassinucellate type. Normally these ovules have fairly large nucellus.
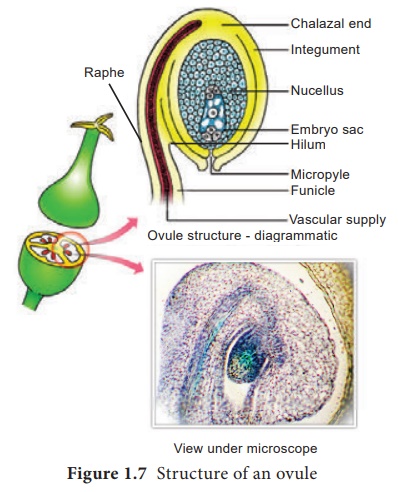
Group of cells found at the base of the ovule between the chalaza and embryo sac is called hypostase
and the thick -walled cells found above the micropylar end above the embryo sac
is called epistase. heT structure of ovule is given in Figure 1.7.
Types of Ovules
The ovules are
classified into six main types based on the orientation, form and position of
the micropyle with respect to funicle and chalaza. Most important ovule types
are orthotropous, anatropous, hemianatropous and campylotropous. The types of
ovule is given in Figure 1.8.
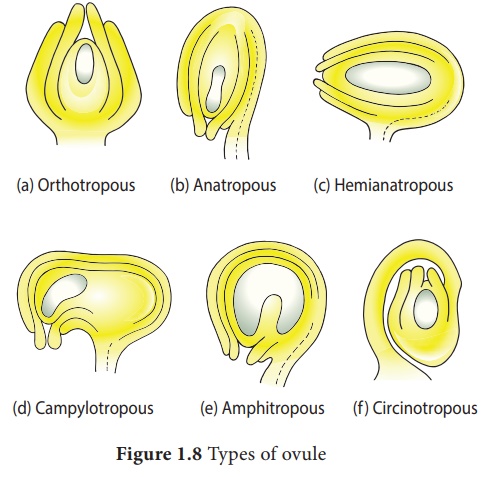
Orthotropous: In this type of ovule,
the micropyle is at the distal end and the micropyle, the funicle and
the chalaza lie in one straight verticalline. Examples: Piperaceae,
Polygonaceae
Anatropous: The body of the ovule
becomes completely inverted so that the micropyle and funiculus come to
lie very close to each other. This is the common type of ovules found in dicots
and monocots.
Hemianatropous: In this, the body of the
ovule is placed transversely and at right angles to the funicle. Example:
Primulaceae.
Campylotropous: The body of the ovule at
the micropylar end is curved and more or less bean shaped. The embryo
sac is slightly curved. All the three, hilum, micropyle and chalaza are
adjacent to one another, with the micropyle oriented towards the placenta.
Example: Leguminosae
In addition to the above main types there are two more types of ovules they are, Amphitropous: The distance between hilum and chalaza is less. The curvature of the ovule leads to horse-shoe shaped nucellus. Example: some Alismataceae.
Circinotropous: Funiculus is very long
and surrounds the ovule. Example: Cactaceae
Megasporogenesis
The process of
development of a megaspore from a megaspore mother cell is called megasporogenesis.
As the ovule develops, a
single hypodermal cell in the nucellus becomes enlarged and functions as archesporium.
In some plants, the archesporial cell may directly function as megaspore mother
cell. In others, it may undergo a transverse division to form outer primary
parietal cell and inner primary sporogenous cell. The parietal cell may remain
undivided or divide by few periclinal and anticlinal divisions to embed the
primary sporogenous cell deep into the nucellus. The primary sporogenous cell
functions as a megaspore mother cell. The megaspore mother cell undergoes
meiotic division to form four haploid megaspores. Based on the number of
megaspores that develop into the Embryo sac, we have three basic types of
development: monosporic, bisporic and tetrasporic. The megaspores
are usually arranged in a linear tetrad. Of the four megaspores formed, usually
the chalazal one is functional and other three megaspores degenerate. The
functional megaspore forms the female gametophyte or embryo sac. This type of
development is called monosporic development (Example: Polygonum).
Of the four megaspores formed if two are involved in Embryo sac formation
the development is called bisporic (Example: Allium). If all the
four megaspores are involved in Embryo sac formation the development is
called tetrasporic (Example: Peperomia). An ovule generally has a
single embryo sac. The development of monosporic embryo sac (Polygonum
type) is given in Figure 1.9.
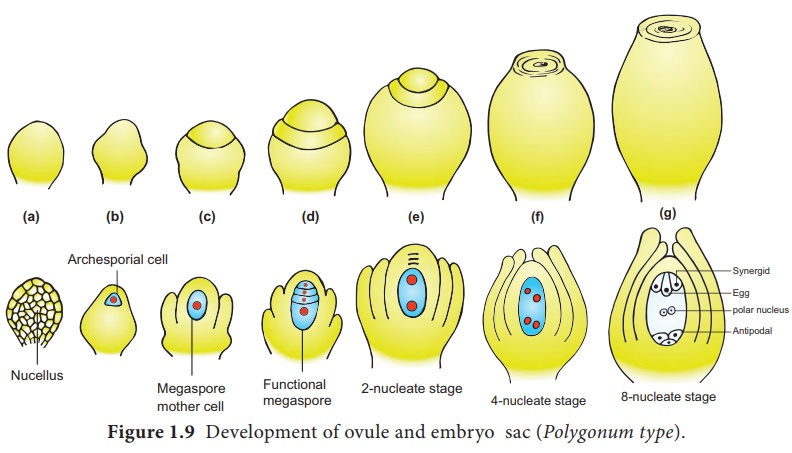
Development of Monosporic embryo sac.
To describe the stages
in embryo sac development and organization the simplest monosporic type of
development is given below.
The functional megaspore is the first cell of the embryo sac or female gametophyte. The megaspore elongates along micropylar-chalazal axis. The nucleus undergoes a mitotic division. Wall formation does not follow the nuclear division. A large central vacuole now appears between the two daughter nuclei. The vacuole expands and pushes the nuclei towards the opposite poles of the embryo sac. Both the nuclei divide twice mitotically, forming four nuclei at each pole. At this stage all the eight nuclei are present in a common cytoplasm (free nuclear division). After the last nuclear division the cell undergoes appreciable elongation , assuming a sac-like appearance.
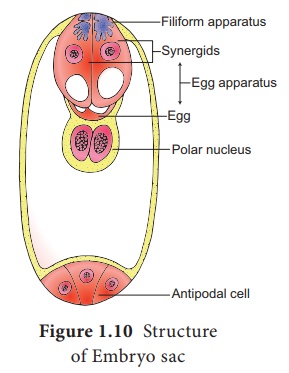
This is followed by cellular organization of the embryo sac. Of the
four nuclei at the micropylar end of the embryo sac, three organize into an egg
apparatus, the fourth one is left free in the cytoplasm of the central cell
as the upper polar nucleus. Three nuclei of the chalazal end form three antipodal
cells whereas the fourth one functions as the lower polar
nucleus. Depending on the plant the 2 polar nuclei may remain free or
may fuse to form a secondary nucleus (central cell). The egg apparatus
is made up of a central egg cell and two synergids, one on each side of the egg
cell. Synergids secrete chemotropic substances that help to attract the pollen
tube. The special cellular thickening called filiform apparatus of synergids
help in the absorption, conduction of nutrients from the nucellus to embryo
sac. It also guides the pollen tube into the egg. Thus, a 7 celled with 8
nucleated embryo sac is formed. The structure of embryo sac is given in Figure
1.10.
Related Topics