Chapter: Biochemistry: Biochemistry and the Organization of Cells
Common Ground for All Cells
Common Ground for All Cells
Did eukaryotes develop from prokaryotes?
The complexity of eukaryotes raises many questions about how such cells arose from simpler progenitors. Symbiosis plays a large role in current theories of the rise of eukaryotes; the symbiotic association between two organisms is seen as giving rise to a new organism that combines characteristics of both the origi-nal ones. The type of symbiosis called mutualism is a relationship that benefits both species involved, as opposed to parasitic symbiosis, in which one species gains at the other’s expense. A classic example of mutualism (although it has been questioned from time to time) is the lichen, which consists of a fungus and an alga. The fungus provides water and protection for the alga; the alga is photosynthetic and provides food for both partners. Another example is the root-nodule system formed by a leguminous plant, such as alfalfa or beans, and anaerobic nitrogen-fixing bacteria (Figure 1.21).
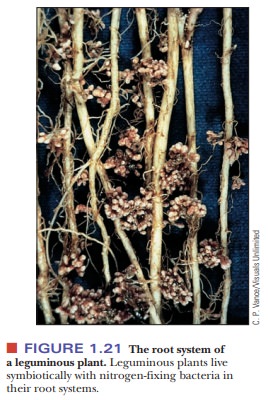
The plant gains useful compounds
of nitrogen, and the bacteria are protected from oxygen, which is harmful to
them. Still another example of mutualistic symbiosis, of great practical
interest, is that between humans and bacteria, such as Escherichia coli, that live in the intestinal tract. The bacteria receive
nutrients and protection from their immediate environment. In return, they aid
our digestive process. Without beneficial intestinal bacteria, we would soon
develop dysentery and other intestinal disorders. These bacteria are also a
source of certain vitamins for us, because they can synthesize these vitamins
and we cannot. The disease-causing strains of E. coli that have been in the news from time to time differ
markedly from the ones that naturally inhabit the intestinal tract.
Did symbiosis play a role in the development of eukaryotes?
In
hereditary symbiosis, a larger host cell contains a genetically determined
number of smaller organisms. An example is the protist Cyanophora paradoxa, a eukaryotic host that contains a genetically
determined number of cyanobacteria (blue-green algae). This relationship is an
example of endosymbiosis, because
the cyanobacteria are contained within the host organism. The cyanobacteria are
aerobic prokaryotes and are capable of photosynthesis (Figure 1.22). The host
cell gains the products of photosynthesis; in return, the cyanobacteria are
protected from the environment and still have access to oxygen and sunlight
because of the host’s small size. In this model, with the passage of many
gen-erations, the cyanobacteria would have gradually lost the ability to exist
inde-pendently and would have become organelles within a new and more complex
type of cell. Such a situation in the past may well have given rise to
chloroplasts, which are not capable of independent existence. Their autonomous
DNA and their apparatus for synthesizing ribosomal proteins can no longer meet
all their needs, but the fact that these organelles have their own DNA and are
capable of protein synthesis suggests that they may have existed as independent
organ-isms in the distant past.

A
similar model can be proposed for the origin of mitochondria. Consider this
scenario: A large anaerobic host cell assimilates a number of smaller aero-bic
bacteria. The larger cell protects the smaller ones and provides them with
nutrients. As in the example we used for the development of chloroplasts, the
smaller cells still have access to oxygen. The larger cell is not itself
capable of aerobic oxidation of nutrients, but some of the end products of its
anaerobic oxidation can be further oxidized by the more efficient aerobic
metabolism of the smaller cells. As a result, the larger cell can get more
energy out of a given amount of food than it could without the bacteria. In
time, the two associated organisms evolve to form a new aerobic organism, which
contains mitochon-dria derived from the original aerobic bacteria.
The fact
that both mitochondria and chloroplasts have their own DNA is an important
piece of biochemical evidence in favor of this model. Additionally, both mitochondria
and chloroplasts have their own apparatus for synthesis of RNA and proteins.
The genetic code in mitochondria differs slightly from that found in the
nucleus, which supports the idea of an independent origin. Thus, the remains of
these systems for synthesis of RNA and protein could reflect the organelles’
former existence as free-living cells. It is reasonable to conclude that large
unicellular organisms that assimilated aerobic bacteria went on to evolve
mitochondria from the bacteria and eventually gave rise to animal cells. Other
types of unicellular organisms assimilated both aerobic bacteria and
cya-nobacteria and evolved both mitochondria and chloroplasts; these organisms
eventually gave rise to green plants.
The proposed connections between prokaryotes and eukaryotes are not established with complete certainty, and they leave a number of questions unanswered. Still, they provide an interesting frame of reference from which to consider evolution and the origins of the reactions that take place in cells.
Summary
Many theories about the rise of eukaryotes from
prokaryotes focus on a possible role for symbiosis.
The idea
of endosymbiosis, in which a larger cell engulfs a smaller one, plays a large
role in scenarios for the development of organelles in eukaryotic cells.
Related Topics