Chapter: Psychology: Personality
The Trait Approach: Traits and Biology
Traits and Biology
Where
does all of this leave us? Plainly, situations do matter in shaping how we act,
and, as a result, we can easily document inconsistencies in how someone
behaves: She might be honest in one setting but treacherous in another,
friendly in one situation but hostile otherwise, with her behavior in each case
governed as much by where she is as by who she is. At the same time, we can
also document ways in which each of us is consistent in who we are. We
shouldn’t be surprised, therefore, that personality traits have been shown to
predict important life outcomes (Ozer & Benet-Martinez, 2006; Roberts,
Kuncel, Shiner, Caspi, & Goldberg, 2007). For example, the Big Five
personality dimensions are related to outcomes ranging from career success
(Barrick, Mount, & Gupta, 2003) to crim-inal activities (Wiebe, 2004) to
health and mortality (Roberts, Walton, & Bogg, 2005).
Personality
consistency thus seems to be alive and well, especially if we understand trait
labels to be descriptions of how a person tends to act in a certain sort of
situation, rather than a description of what he is like at all times and in all
places. However, this simply leads to a new question: Given that people do
differ in their personalities, how do these differences arise?
GENES
AND
PERSONALITY
Mounting
evidence suggests that personality traits grow out of the individual’s temperament, a characteristic pattern
of emotion, attention, and behavior that isevident from an early age and is
determined to a considerable degree by genetic pat-terns (Kagan, 1994; Rothbart
& Bates, 2006; Thomas & Chess, 1977). Evidence for this genetic
influence—both on temperament and on personality in general—comes from the same
methods used to study heritability in other contexts, including stud-ies of twins
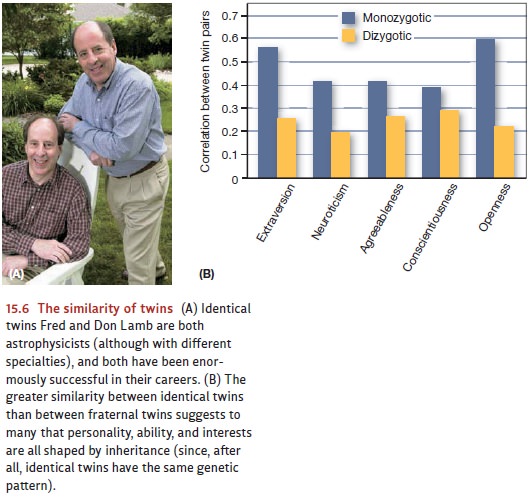
(monozygous or not, raised separately or apart) and studies of adoptees (Figure
15.6). Data from these studies tell us that in just about all cases, identical
twins turn out to be more alike than fraternal twins on various personality
attributes (see A. H. Buss & Plomin, 1984; Zuckerman, 1987a). For example,
one study com-pared the personalities of 123 pairs of identical twins and 127
pairs of fraternal twins and found that heritability for the Big Five
personality dimensions ranged from 40 to 60% (Borkenau, Riemann, Angleitner,
& Spinath, 2001; Jang, Livesley, & Vernon, 1996; Loehlin, 1992).
Peculiarly, though, genetic influences have also been identified for much more specific traits such as television watching, traditionalism, and the willingness to divorce (Bouchard, Lykken, McGue, Segel, & Tellegen, 1990; McGue & Lykken, 1992; Plomin, Corley, DeFries, & Fulker, 1990). In each case, there is a greater resemblance between identical twins with regard to these traits than between fraternal twins. Of course, natural selection unfolds at a slow pace, but television has existed for less than a century. Therefore, the genetic influence on these kinds of tendencies may reflect the operation of other, more general personality dimensions—such as those named in the Big Five. For example, television watching may be associated with extraversion, and traditionalism with conscientiousness. Likewise, in a study of adult twins, those twins who divorced had higher average scores on measures related to extraversion and neuroticism, and lower scores on measures of impulse control (Jockin, McGue, & Lykken, 1996). This is certainly consistent with the idea that the specific trait (tendency to divorce) is derived from the more general biologically based tendencies such as extraversion or neuroticism, and it is the latter that are influenced by the genes.
PHYSIOLOGY AND PERSONALITY
If
our personalities have a basis in our genes, what exactly do the genes code
for? Using physiological data, psychologists
are beginning to explore how people with differentpersonality traits differ in
their biological functioning, with the hope that these data will offer a
glimpse into how our genes shape who we are.
One
example of this work is inspired by Hans Eysenck’s theory of extraversion/
introversion. Eysenck proposed that the observable difference in personality
derives from the fact that introverts react more strongly than extraverts to
external stimuli (M. W. Eysenck, 1987). As a result, he argued, introverts
often guard against stimula-tion from the outside, which to them feels like
overstimulation. This is why, for exam-ple, introverts shy away from social
settings—the stimulation in that setting would be more than they are
comfortable with. It also turns out that introverts have a lower tol-erance for
pain (Bartol & Costello, 1976), and, when they are studying, they prefer
less noise and fewer opportunities for socializing (J. B. Campbell &
Hawley, 1982).
Can
we confirm this proposal through studies of the brain? In one study,
investiga-tors measured how people’s brain stems reacted when the people heard
clicking noises. In line with Eysenck’s theory, introverts showed a quicker
response than extraverts, indicating more reactive brain stems (Bullock &
Gilliland, 1993; also see Kumari, Ffytche, Williams, & Gray, 2004).
Similar
arguments may help us to understand a more specific trait, sensationseeking, which refers to the tendency to seek varied and
novel experiences, to look forthrills and adventure, and to be highly
susceptible to boredom (Zuckerman, 1979, 1994; Figure 15.7). People high in
sensation-seeking are more likely to participate in risky sports; to get more
restless in monotonous, confined situations; and to drive faster than people
with low sensation-seeking scores (Zuckerman, 1979, 1983).

What leads to this pattern of behavior? One suggestion is that sensation seekers are people whose neurotransmitter systems (especially those relying on norepinephrine and dopamine) are underreactive. As a result, these people are chronically underaroused, and this makes them seek thrills and take risks to jog their sluggish neurotransmitter systems into greater activity (Zuckerman, 1987b, 1990, 1994). Consistent with this hypothesis, sensation seekers seem to be at greater risk for abusing drugs that influence dopamine lev-els. Researchers explain this finding by suggesting that in this case, the sensation seekers are using drugs, rather than activities such as sky diving or snake handling, to activate their underactive brain systems (Bardo, Donohew, & Harrington, 1996). These findings are buttressed by those of researchers who have developed a “rat model” of drug use.
Using
this model, researchers have shown that high levels of sensation seeking
(measured by animals’ activity levels in a novel environment) predict the
animals’ propensity to give themselves cocaine (Belin, Mar, Dalley, Robbins,
& Everitt, 2008).
Sensation
seeking is common among extraverts; a very different pattern—called inhibited temperament—is associated
with introversion and neuroticism (Fox,Henderson, Marshall, Nichols, &
Ghera, 2005; Kagan, 1994, 2003; Kagan & Snidman, 1991; Putnam & Stifter,
2005). As infants, people with inhibited temperaments tend to react strongly
when they are distressed, crying vigorously and showing high levels of motor
activity. As young children, inhibited individuals are unwilling to approach
novel stimuli or people, become anxious in new situations, and frequently seek
reassurance from their caregivers. Adolescents and adults who were categorized
as inhibited at an early age are much less likely than others to be outgoing
and socially spontaneous.
The
explanation for inhibited temperament may mirror the account of sensation
seeking, and in particular may be associated with an overreactive brain (and
attempts to compensate for this) just as sensation seeking is associated with
an underreactive brain. Specifically, Kagan and colleagues hypothesize that
inhibited children have a low threshold for activity in the amygdala, a
subcortical brain structure crucial for detecting important stimuli in the
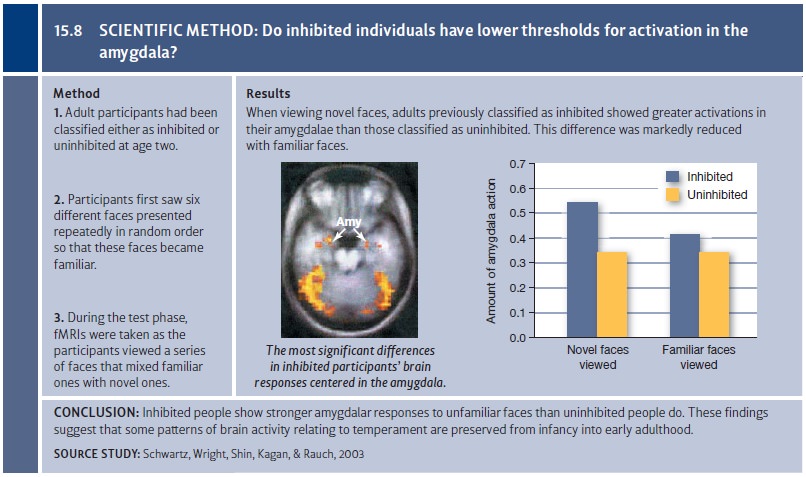
environment. This hypothesis was tested using an fMRI study of young adults who
had been classified when they were infants as either inhibited or uninhibited
(Schwartz, Wright, Shin, Kagan, & Rauch, 2003; Figure 15.8). These
participants viewed a series of familiar and unfamiliar faces. As predicted,
when the inhibited participants viewed unfamiliar faces, their amygdalae showed
higher levels of activation than did the amygdalae of uninhibited participants.
The two groups did not differ, however, when they were viewing familiar faces,
suggesting that it was the newness of the faces that led the inhibited
individuals’ amygdalae to respond more strongly.
Related Topics