Chapter: Human Neuroanatomy(Fundamental and Clinical): Introduction to Neuroanatomy
The Synapse - Human Neuroanatomy
The Synapse
We have seen that synapses are sites of junction between neurons. Synapses may be of various types depending upon the parts of the neurons that come in contact. In the most common type of synapse, an axon terminal establishes contact with the dendrite of a receiving neuron to form an axodendritic synapse. Synapses on dendrites may be located on spines or on the smooth areas between spines. The axon terminal may synapse with the cell body (axosomatic synapse) or, less commonly, with the axon of the receiving neuron (axoaxonal synapse). An axoaxonal synapse may be located either on the initial segment (of the receiving axon) or just proximal to an axon terminal.
In some parts of the brain (e.g., the thalamus) we see some synapses in which the presynaptic element is a dendrite instead of an axon. Such synapses may be dendro-axonic or dendro-dendritic. In yet others the soma of a neuron may synapse with the soma of another neuron (somato-somatic synapse), or with a dendrite (somato-dendritic synapse).
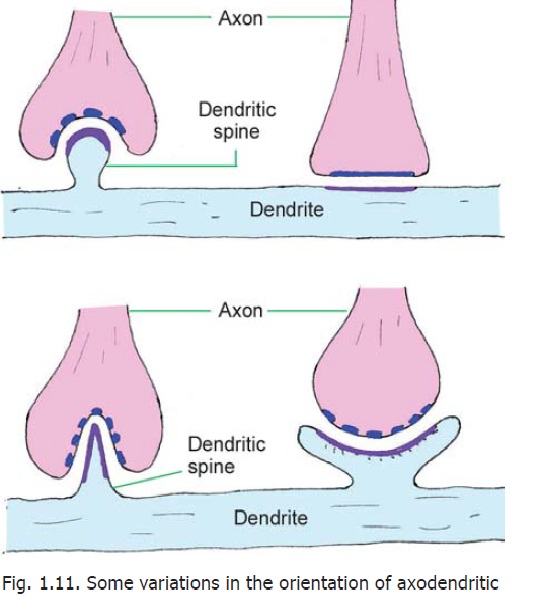
The axon may terminate in a single bulb-like end called a bouton (or synaptic bag). Alternatively, the terminal part of the axon may bear a number of such enlargements each of which synapses with the receiving neuron. We have seen that dendrites bear numerous spines. Axon terminals may synapse either with the spines or with smooth portions of the dendrite between the spines. Occasionally, an axon terminal may end by synapsing with the terminal bouton of another axon forming what is called a serial synapse. In certain situations several neurons may take part in forming complex synapses. Such areas, encapsulated by neuroglial cells, form synaptic glomeruli. Such glomeruli are found in the cerebellum, the olfactory bulb, the lateral geniculate body and in some other situations.
At some sites several synapses may be present around a short length of a dendrite and may be enclosed within a glial capsule. Such a complex is called a synaptic cartridge.
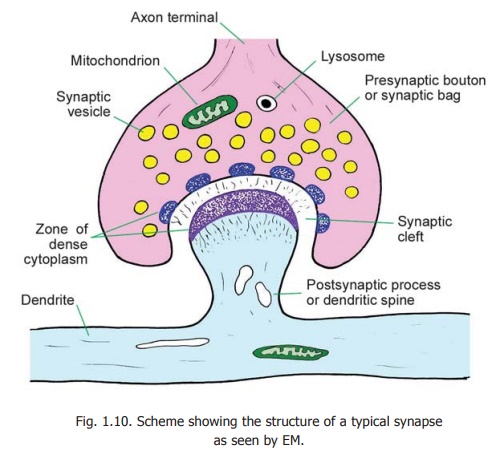
A synapse transmits an impulse only in one direction. The two elements taking part in a synapse can, therefore, be spoken of aspresynaptic and postsynaptic (Fig. 1.10). In an axo-dendritic synapse, the terminal enlargement of the axon may be referred to as thepresynaptic bouton or synaptic bag.The region of the dendritereceiving the axon terminal is the postsynaptic process. The two areseparated by a space called the synapticcleft.Delicate fibres or granular materialmay be seen within the cleft. On either side of the cleft there is a region of dense cytoplasm. On the presynaptic side this dense cytoplasm is broken up into several bits. On the postsynaptic side the dense cytoplasm is continuous and is associated with a meshwork of filaments called the synaptic web.
The thickened areas of membrane on the presynaptic and postsynaptic sides constitute the active zone of a synapse. Neuro-transmission takes place through this region. Some variations in the structure of the active zone are described below.
Within the presynaptic bouton numerous synaptic vesicles can be seen. Mitochondria and lysosomes may also be present. The presynaptic bouton contains numerous microtubules (that extend into it from the axon). The tubules end near the presynaptic membrane. Synaptic vesicles are attached to the microtubules by short stalks. The postsynaptic process may also show membranous structures of various shapes, microtubules, filaments and endoplasmic reticulum.
Various proteins and enzymes are present in relation to presynaptic and postsynaptic regions. Some of them (F-actin, spectrin) form a filamentous network that immobilises vesicles until they are to be released.
Neurotransmitters
The transmission of impulses through synapses involves the release of chemical substances called neurotransmitters that are present within synaptic vesicles. When a nerve impulse reaches a terminalbouton neurotransmitter is released into the synaptic cleft. Under the influence of the neurotransmitter the postsynaptic surface becomes depolarised resulting in a nerve impulse in the postsynaptic neuron. In the case of inhibitory synapses, the presence of the neurotransmitter causes hyperpolarisation of the postsynaptic membrane. The neurotransmitter released into the synaptic cleft acts only for a very short duration. It is either destroyed (by enzymes) or is withdrawn into the terminal bouton.
When an action potential reaches the presynaptic terminal, voltage sensitive calcium channels are opened up so that there is an influx of calcium ions leading to a series of chemical changes. As a result of these changes synaptic vesicles pour the neurotransmitter stored in them into the synaptic cleft. The neurotransmitter reaches and binds onto receptor molecules present in the postsynaptic membrane. This alters permeability of the postsynaptic membrane to ions of calcium, sodium, potassium or chloride leading to depolarisation (or hyperpolarisation at inhibitory synapses). The best known (or classical) neurotransmitters responsible for fast but short-lived action of the kind described above are acetylcholine, noradrenaline and adrenaline. For long, all nerve terminals were regarded as either cholinergic or adrenergic, until it was recognised that these were not the only neurotransmitters present. Other fast neurotransmitters whose presence is now well established are dopamine and histamine.
It is also recognised that apart from the neurotransmitters mentioned above numerous other chemical substances are associated with synapses. Some of these, which probably act as neuro-transmitters, are serotonin, gama-aminobutyric acid (GABA), glutamate, aspartate and glycine.
It is now known that at some synapses the effect of a neurotransmitter may last for seconds or even minutes. Repeated synaptic activity can have long lasting effects on the receptor neuron including structural changes such as the formation of new synapses, alterations in the dendritic tree, or growth of axons. Such effects produced under the influence of chemical substances are described asneuromediation, the chemical substances concerned being called neuromediators. This term includes neurohormones, synthesised in neurons and poured into the blood stream through terminals resembling synapses in structure. Similar chemical substances are also poured into the cerebrospinal fluid or into intercellular spaces to influence other neurons in a diffuse manner.
Lastly, some chemical substances associated with synapses do not influence synaptic transmission directly, but influence the effects of transmitters or of neuromediators. Such chemical substances are referred to as neuromodulators. Several peptides found in the nervous system probably act as neuromodulators. These include substance P, vasoactive intestinal polypeptide (VIP), somatostatin, cholecystokinin and many others.
The following factors can influence synaptic transmission (and thereby the speed of responses).
a. Drugs like caffeine produce their stimulatory effect by stimulating synaptic transmission.
b. Synaptic transmission may decrease in old age because calcium ion channels become fewer. In the case of the heart this may impair the stimulating effect of exercise on heart rate and cardiac output.
c. Synaptic transmission is disturbed in some diseases like myasthenia gravis.
d. It is also affected in poisoning by organophosphates. In this condition the action of acetylcholine esterase is inhibited and acetyl choline accumulates. This can lead to spasm of respiratory muscles and death.
Classification of Synapses
Synapses may be of various types depending on the neuronal elements taking part. They may also be classified on the basis of their ultrastructure, and on the basis of the neurotransmitters released by them. From a physiological standpoint a synapse may be excitatory or inhibitory. Synapses in different situations can vary considerably in overall shape (Fig. 1.11); in the size, shape and nature of synaptic vesicles and in the configuration of the presynaptic and postsynaptic areas of dense cytoplasm.
Two main types of synapses are recognised on the basis of their ultrastructure.
Asymmetric or Type I synapses:In these synapses the subsynaptic zone of dense cytoplasm isthicker on the presynaptic side. The synaptic cleft is about 30 nm. Such synapses are excitatory.
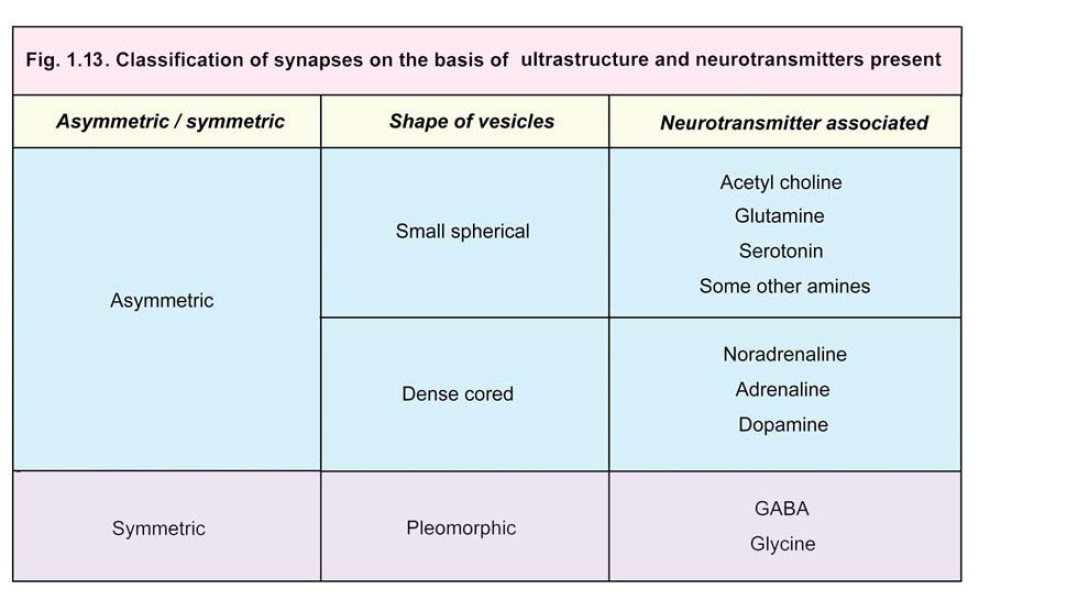
Symmetric or Type II synapses:In these synapses the subsynaptic zones of dense cytoplasm arethin and of similar thickness on both sides. The synaptic cleft measures about 20 nm.Various synapses intermediate in structure between these two main types are also encountered. The vesicles to be seen within synapses can also be of various types. Some vesicles are clear while others have dense cores. They may be pleomorphic (i.e., a mixture of various shapes). The appearance of vesicles can often be correlated with the neurotransmitter present. On the basis of these characters some sub-varieties of Type 1 and Type II synapses that have been recognised are given in Fig. 1.13.
Through its ramifications an axon usually establishes synapses with several different neurons; but in some situations it synapses with one neuron only. Some axons bear boutons that do not come into direct contact with other neurons. Such boutons may represent areas where neurotransmitters are released into surrounding areas, and can have widespread rather than localised effect.
At some sites specialised regions may be seen in relation to synapses. They are mentioned here as they are given specific names.
In some synapses in the retina and internal ear vesicles are arranged around a rod-like element placed at right angles to the cell membrane. This configuration is called a synaptic ribbon. Within some dendritic spines collections of flattened cisternae (endoplasmic reticulum) with associated dense material are seen. These are given the name spine apparatus.
Electrical Synapses
Synapses involving the release of neurotransmitters are referred to as chemical synapses. At some sites one cell may excite another without the release of a transmitter. At such sites adjacent cells have direct channels of communication through which ions can pass from one cell to another altering their electrical status. Such synapses are called electrical synapses.
At the site of an electrical synapse plasma membranes (of the two elements taking part) are closely applied, the gap between them being about 4 nm. Proteins called connexins project into this gap from the membrane on either side of the synapse. The proteins are so arranged that small open channels are created between the two synaptic elements.
Electrical synapses are common in lower vertebrates and invertebrates. They have been demonstrated at some sites in the brains of mammals (e.g., in the inferior olive and cerebellum).
Junctions between receptors and neurons, or between neurons and effectors, share some of the features of typical synapses and may also be regarded as synapses. Junctions between cardiac myocytes, and between smooth muscle cells, are regarded as electrical synapses.
Influence of neural activity on synapses
It has been shown that neural activity acts as a stimulus for development of new synapses and for increase in their size, specially in early postnatal life. Some experiments show that even in later life (in some situations) brief synaptic activity can have an influence on the subsequent activity of the synapse. This is specially true in areas like the hippocampus and may be associated with memory.
Related Topics