Chapter: Human Neuroanatomy(Fundamental and Clinical): Introduction to Neuroanatomy
Neuron Structure
Neuron Structure
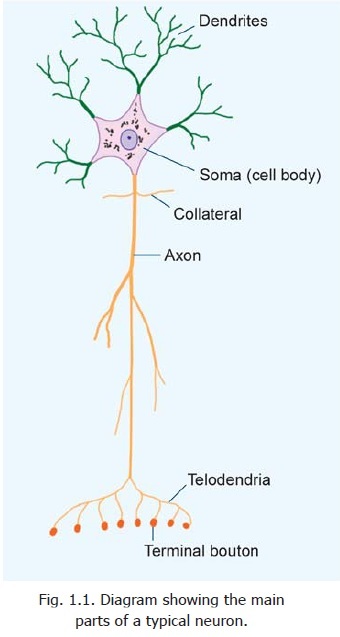
Elementary Structure of a Typical Neuron
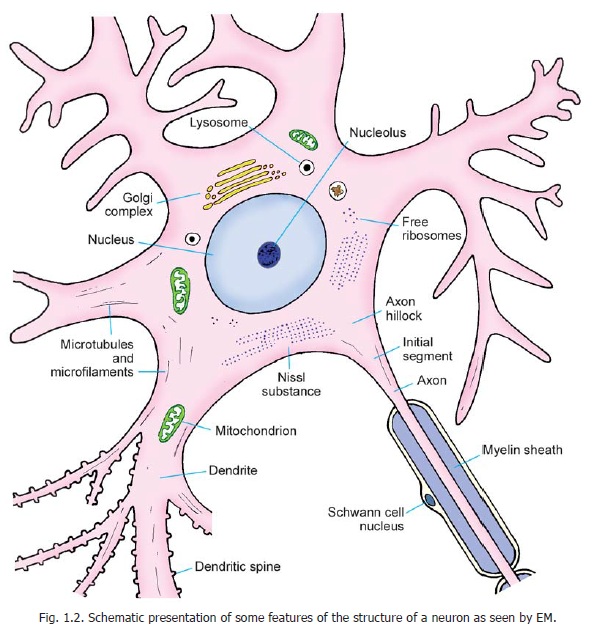
Neurons vary considerably in size, shape and other features. However, most of them have some major features in common and these are described below (Figs. 1.1 to 1.4).
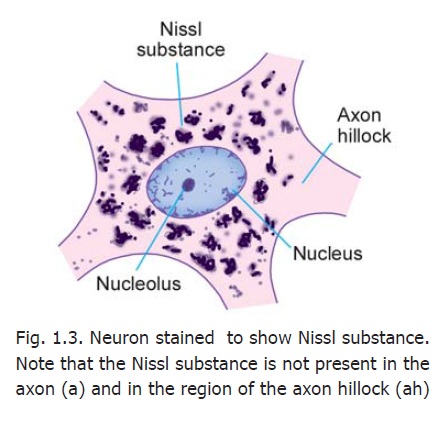
A neuron consists of a cell body that gives off a number of processes. The cell body is also called the soma or perikaryon. Like a typical cell it consists of a mass of cytoplasm surrounded by a cell membrane. The cytoplasm contains a large central nucleus (usually with a prominent nucleolus), numerous mitochondria, lysosomes and a Golgi complex (Fig. 1.2). In the past it has often been stated that centrioles are not present in neurons, but studies with the electron microscope (usually abbreviated to EM) have shown that centrioles are present. In addition to these features, the cytoplasm of a neuron has some distinctive characteristics not seen in other cells. The cytoplasm shows the presence of a granular material that stains intensely with basic dyes; this material is the Nissl substance (also called Nissl bodies or granules) (Fig. 1.3). When examined with the EM, these bodies are seen to be composed of rough surfaced endoplasmic reticulum (Fig 1.2). The presence of abundant granular endoplasmic reticulum is an indication of the high level of protein synthesis in neurons. The proteins are needed for maintenance and repair, and for production of neurotransmitters and enzymes.
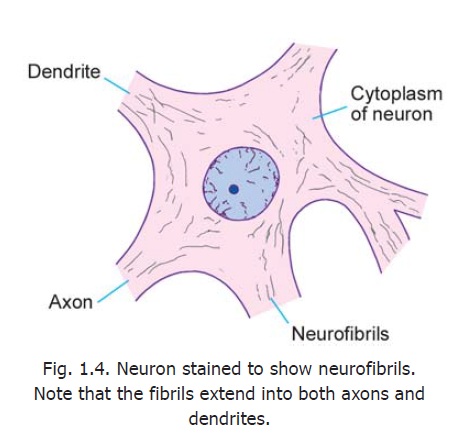
Another distinctive feature of neurons is the presence of a network of fibrils permeating the cytoplasm (Fig. 1.4). Theseneurofibrils are seen, with the EM, to consist of microfilaments and microtubules. (The centrioles present in neurons may be concerned with the production and maintenance of microtubules).
Some neurons contain pigment granules (e.g., neuromelanin in neurons of the substantia nigra). Ageing neurons contain a pigment lipofuscin (made up of residual bodies derived from lysosomes).
The processes arising from the cell body of a neuron are called neurites. These are of two kinds. Most neurons give off a number of short branching processes called dendrites and one longer process called an axon.
The dendrites are characterised by the fact that they terminate near the cell body. They are irregular in thickness, and Nissl granules extend into them. They bear numerous small spines that are of variable shape.
The axon may extend for a considerable distance away from the cell body. The longest axons may be as much as a metre long. Each axon has a uniform diameter, and is devoid of Nissl substance.
In addition to these differences in structure, there is a fundamental functional difference between dendrites and axons. In a dendrite, the nerve impulse travels towards the cellbody whereas in an axon the impulse travels away from the cell body.
We have seen above that the axon is free of Nissl granules. The Nissl-free zone extends for a short distance into the cell body: this part of the cell body is called the axon hillock. The part of the axon just beyond the axon hillock is called the initial segment(Fig. 1.2).
During its formation each axon comes to be associated with certain cells that provide a sheath for it. The cells providing this sheath for axons lying outside the central nervous system are called Schwann cells. Axons lying within the central nervous system are provided a similar covering by a kind of neuroglial cell called an oligodendrocyte. The nature of this sheath isbest understood by considering the mode of its formation (Fig. 1.5). An axon lying near a Schwann cell (1) invaginates into the cytoplasm of the Schwann cell (2,3). In this process the axon comes to be suspended by a fold of the cell membrane of the Schwann cell: this fold is called the mesaxon (3). In some situations the mesaxon becomes greatly elongated and comes to be spirally wound around the axon, which is thus surrounded by several layers of cell membrane (4,5). Lipids are deposited between adjacent layers of the membrane. These layers of the mesaxon, along with the lipids, form the myelin sheath. Outside the myelin sheath athin layer of Schwann cell cytoplasm persists to form an additional sheath which is called the neurilemma (also called the neurilemmalsheath or Schwann cell sheath). Axons having
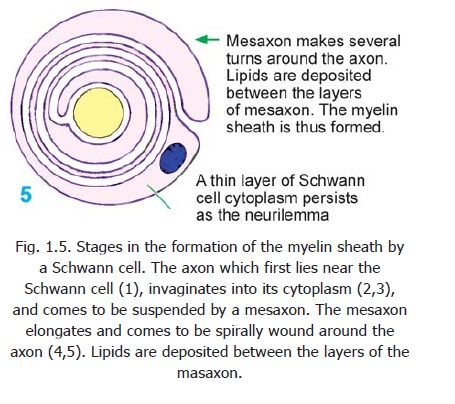
a myelin sheath are called myelinated axons. The presence of a myelin sheath increases the velocity of conduction (for a nerve fibre of the same diameter). It also reduces the energy expended in the process of conduction.
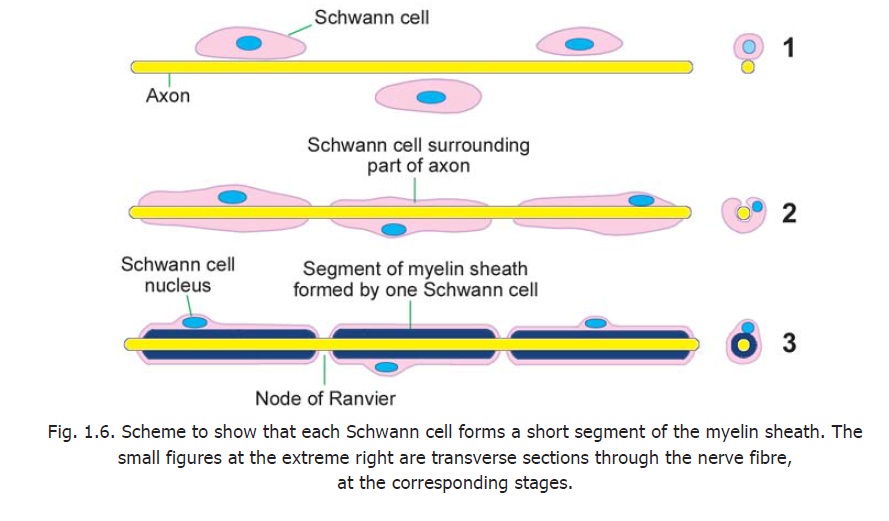
An axon is related to a large number of Schwann cells over its length (Fig. 1.6). Each Schwann cell provides the myelin sheath for a short segment of the axon. At the junction of any two such segments there is a short gap in the myelin sheath. These gaps are called the nodes of Ranvier.
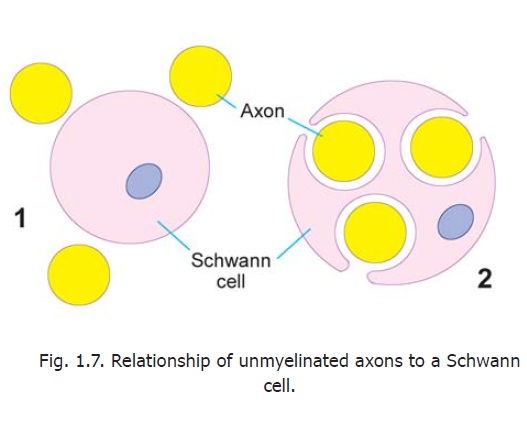
There are some axons that are devoid of myelin sheaths. These unmyelinated axons invaginate into the cytoplasm of Schwann cells, but the mesaxon does not spiral around them (Fig. 1.7). Another difference is that several such axons may invaginate into the cytoplasm of a single Schwann cell.
An axon may give off a variable number of branches (Fig. 1.1). Some branches, that arise near the cell body and lie at right angles to the axon are called collaterals. At its termination the axon breaks up into a number of fine branches called telodendria that may end in small swellings (terminalboutons or bouton terminaux). An axon (or its branches) can terminate in two ways. Within thecentral nervous system, it always terminates by coming in intimate relationship with another neuron, the junction between the two neurons being called a synapse. Outside the central nervous system, the axon may end in relation to an effector organ (e.g., muscle or gland), or may end by synapsing with neurons in a peripheral ganglion.
Axons (and some dendrites that resemble axons in structure:) constitute what are commonly called nerve fibres.
Grey and White Matter
Sections through the spinal cord or through any part of the brain show certain regions that appear whitish, and others that have a darker greyish colour. These constitute the white and grey matter respectively. Microscopic examination shows that the cell bodies of neurons are located only in grey matter that also contains dendrites and axons starting from or ending on the cell bodies. Most of the fibres within the grey matter are unmyelinated. On the other hand the white matter con-sists predominantly of myelinated fibres. It is the reflection of light by myelin that gives this region its whitish appearance. Neuroglia and blood vessels are present in both grey and white matter.
The arrangement of the grey and white matter differs at different situations in the brain and spinal cord. In the spinal cord and brainstem the white matter is on the outside whereas the grey matter forms one or more masses embedded within the white matter (Fig. 2.1). In the cerebrum and cerebellum there is an extensive, but thin, layer of grey matter on the surface. This layer is called thecortex. Deep to the cortex there is white matter, but within the latter several isolated masses of grey matter are present (Fig. 8.9). Such isolated masses of grey matter present any-where in the central nervous system are referred to as nuclei. As grey matter is made of cell bodies of neurons (and the processes arising from or terminating on them) nuclei can be defined as groups of cell bodies of neurons. Aggregations of the cell bodies of neurons may also be found outside the central nervous system. Such aggregations are referred to as ganglia. Some neurons are located in nerve plexuses present in close relationship to someviscera. These are, therefore, referred to as ganglionated plexuses.
The axons arising in one mass of grey matter terminate very frequently by synapsing with neurons in other masses of grey matter. The axons connecting two (or more) masses of grey matter are frequently numerous enough to form recognisable bundles. Such aggregations of fibres are called tracts. Larger collections of fibres are also referred to as funiculi, fasciculi or lemnisci. (A lemniscusis a ribbon like band). Large bundles of fibres connecting the cerebral or cerebellar hemispheres to the brainstem are calledpeduncles.
Aggregations of processes of neurons outside the central nervous system constitute peripheral nerves.
Neuropil
Many regions of the brain and spinal cord are occupied by a complex meshwork of axon terminals, dendrites and processes of neuroglial cells. This meshwork is called the neuropil.
Variability in Neuron Structure
Variation in the shape of neuronal cell bodies
Neurons vary considerably in the size and shape of their cell bodies (somata) and in the length and manner of branching of their processes. The cell body varies in diameter from about 5 μm, in the smallest neurons, to as much as 120 μm in the largest ones. The shape of the cell body is dependent on the number of processes arising from it. The most common type of neuron gives off several processes and the cell body is, therefore, multipolar (Fig. 1.8). Some neurons have only one axon and one dendrite and are bipolar.
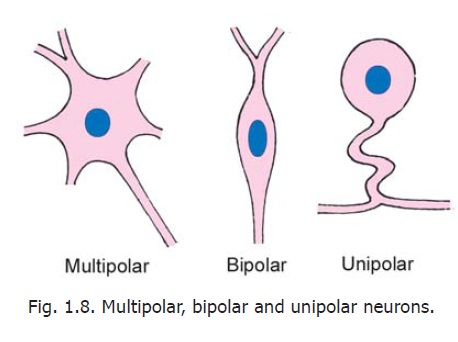
Another type of neuron has a single process (which is highly convoluted). After a very short course this process divides into two. One of the divisions represents the axon; the other is functionally a dendrite, but its structure is indistinguishable from that of an axon. This neuron is described as unipolar, but from a functional point of view it is to be regarded as bipolar. (To avoid confusion on this account this kind of neuron has been referred to, in the past, as a pseudounipolar neuron but this term has now been discarded). Depending on the shapes of their cell bodies some neurons are referred to as stellate (star shaped) or pyramidal.
In addition to the variations in size and shape, the cell bodies of neurons may show striking variations in the appearance of the Nissl substance. In some neurons, the Nissl substance is very prominent and is in the form of large clumps. In some others, the granules are fine and uniformly distributed in the cytoplasm, while yet other neurons show gradations between these extremes. These differences are correlated with function.
Variations in axons
The length of the axon arising from the cell body of a neuron is also subject to considerable variability. Some neurons have long axons, and connect remote regions. These are called Golgi type I neurons. In other neurons axons are short and end near the cell body. They are called Golgi type II neurons or microneurons: these are often inhibitory in function. Very rarely, a neuron may not have a true axon.
As stated earlier, axons also differ in the nature of the sheaths covering them, some of them being myelinated and others unmyelinated. Axons also show considerable variation in the diameter of their cross sections.
Variations in dendrites
Dendrites arising from a neuronal cell body vary considerably in number, and in the extent and manner of branching. They also differ in the distribution of spines on them. These characteristics are of functional importance. The area occupied by the dendrites of a neuron is referred to as its dendriticfield. Different kinds of neurons have differing dendritic fields.
Neurons also show considerable variation in the number and nature of synapses established by them.
FURTHER DETAILS ABOUT NEURONS
Axon hillock and initial segment:
The axon hillock and the initial segment of the axon are of special functional significance. This is the region where action potentials are generated (spike generation) resulting in conduction along the axon. The initial segment is unmyelinated. It often receives axo-axonal synapses that are inhibitory. The plasma membrane here is rich in voltage sensitive channels.
Axoplasmic flow:
The cytoplasm of neurons is in constant motion. Movement of various materials occurs through axons. This axoplasmic flowtakes place both away from and towards the cell body. The flow away from the cell body is greater. Some materials travel slowly (0.1 to 2 mm a day) constituting a slowtransport.In contrast other materials (mainly in the form of vesicles) travel 100 to 400 mm a dayconstituting a rapid transport.
Slow transport is unidirectional, away from the cell body. It is responsible for flow of axoplasm (containing various proteins) down the axon. Rapid transport is bi-directional and carries vesicular material and mitochondria. Microtubules play an important role in this form of transport. Retrograde axoplasmic flow may carry neurotropic viruses along the axon into the neuronal cell body.
Axoplasmic transport of tracer substances introduced experimentally can help to trace neuronal connections.
Role of axoplasmic transport in spread of disease
Some infections that affect the nervous system travel along nerves.
a Rabies is a disease (often fatal) caused by a bite of a rabid dog (and some other animals including monkeys). The saliva of an infected animal contains the rabies virus. The virus travels from the site of the bite to the central nervous system, along nerves, by reverse axoplasmic flow and causes infection there. As this means of transport is slow there is a delay of a few days between the bite and appearance of symptoms. The duration of this delay depends on the length of the nerve fibres concerned. A bite on the face produces symptoms much faster than one on the foot.
b The virus of poliomyelitis is also transported (from the gastrointestinal tract) to the nervous system through reverse axoplasmic flow.
c In contrast tetanus travels from the site of infection to the brain through spaces in the endoneurium of nerve fibres.
Some features of dendrites
a. Dendrites can be distinguished immuno-cytochemically from axons because of the presence in them of microtubule associated protein MAP-2 not present in axons.
b. Dendritic spines vary in size and shape. Some spines contain aggregations of smooth endoplasmic reticulum (in the form of flattened cisternae with associated dense material). The complex is referred to as the spine apparatus.
c. Actin filaments are present in dendritic spines.
d. Some variations in the dendritic field are as follows. The field may be spherical (as in stellate cells), hemispherical,disc-like,conical or flat. In some neurons (e.g., pyramidal), the neuron may have two separate dendritic fields. Apart from shape there is considerable variability in extent of the dendritic field. Some neurons (e.g., Golgi neurons of the cerebellum) have dendritic fields covering a very wide area. More than eighty per cent of the neuronal surface area (excluding the axon) may be situated on the dendritic tree. The frequency of branching of dendrites is correlated with the number of synapses on them. In some neurons the dendritic spines may number several thousand. Finally, it may be emphasised that the dendritic tree is not a ‘fixed’ entity, but may undergo continuous remodelling. This affords a basis for modification of neuronal behaviour.
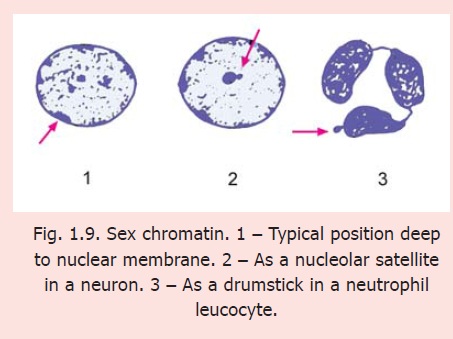
Sex chromatin in neurons
Each cell of a human male has 44+X+Y chromosomes; and each cell of a female has 44+X+X chromosomes. Of the two X-chromosomes in a female only one is functionally active. The other (inactive) X-chromosome forms a mass of dark staining hetero-chromatin that can be identified in suitable preparations and can be useful in determining whether a particular tissue belongs to a male or a female. Because of this association with sex this mass of hetero-chromatin is called the sexchromatin. It is also called a Barr body after the name of the scientist who discovered it. In most cells the sex chromatin lies just under the nuclear membrane (Fig. 1.9). In some cells the sex chromatin occupies a different position from that described above. In neurons it forms a rounded mass lying very close to the nucleolus and is, therefore, called a nucleolar satellite. In neutrophil leucocytes it may appear as an isolated round mass attached to the rest of the nucleus by a narrow band, thus resembling the appearance of a drumstick.
Related Topics