Chapter: 11th Botany : Chapter 14 : Respiration
Stages of Respiration in Plants
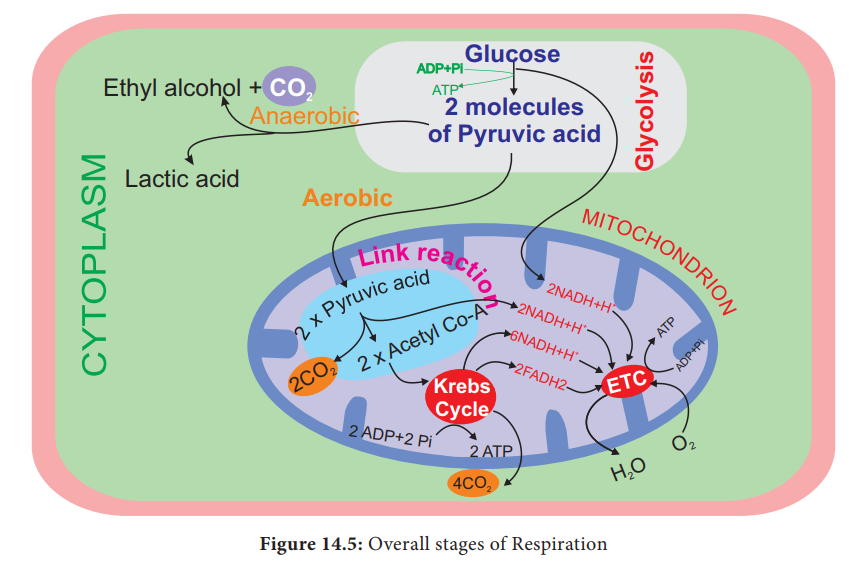
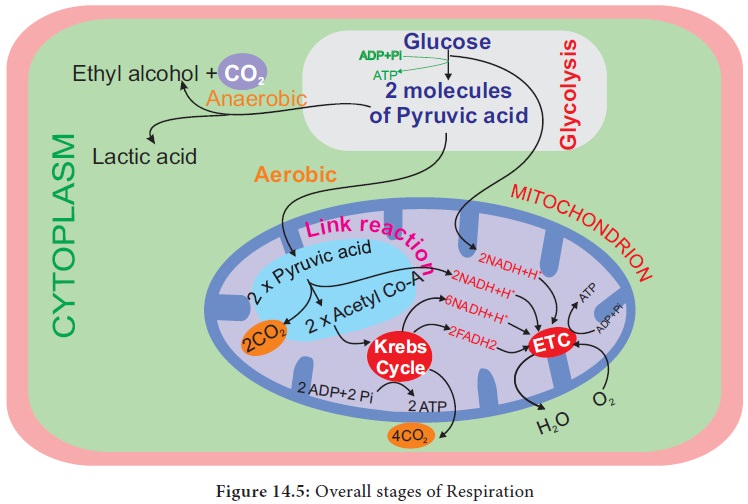
Stages of Respiration
1. Glycolysis-conversion
of glucose into pyruvic acid in cytoplasm of cell.
2. Link
reaction-conversion of pyruvic acid into acetyl coenzyme-A in mitochondrial
matrix.
3. Krebs
cycle-conversion of acetyl coenzyme A into carbon dioxide and water in the
mitochondrial matrix.
4.
Electron transport chain and oxidative phosphorylation remove hydrogen atoms
from the products of glycolysis, link reaction and Krebs cycle release water
molecule with energy in the form of ATP in mitochondrial inner membrane (Figure
14.5).
1. Glycolysis
(Gr: Glykos 5 Glucose, Lysis 5 Splitting) Glycolysis is a
linear series of reactions in which 6-carbon glucose is split into two
molecules of 3-carbon pyruvic acid. The enzymes which are required for
glycolysis are present in the cytoplasm (Figure 14.6). The reactions of
glycolysis were worked out in yeast cells by three scientists Gustav Embden (German), Otto Meyerhoff
(German) and J Parnas (Polish)
and so it is also called as EMP pathway.
It is the first and common stage for both aerobic and anaerobic respiration. It
is divided into two phases.
1.
Preparatory
phase or endergonic phase or hexose
phase (steps 1-5).
2.
Pay off
phase or oxidative phase or exergonic
phase or triose phase (steps 6-10).
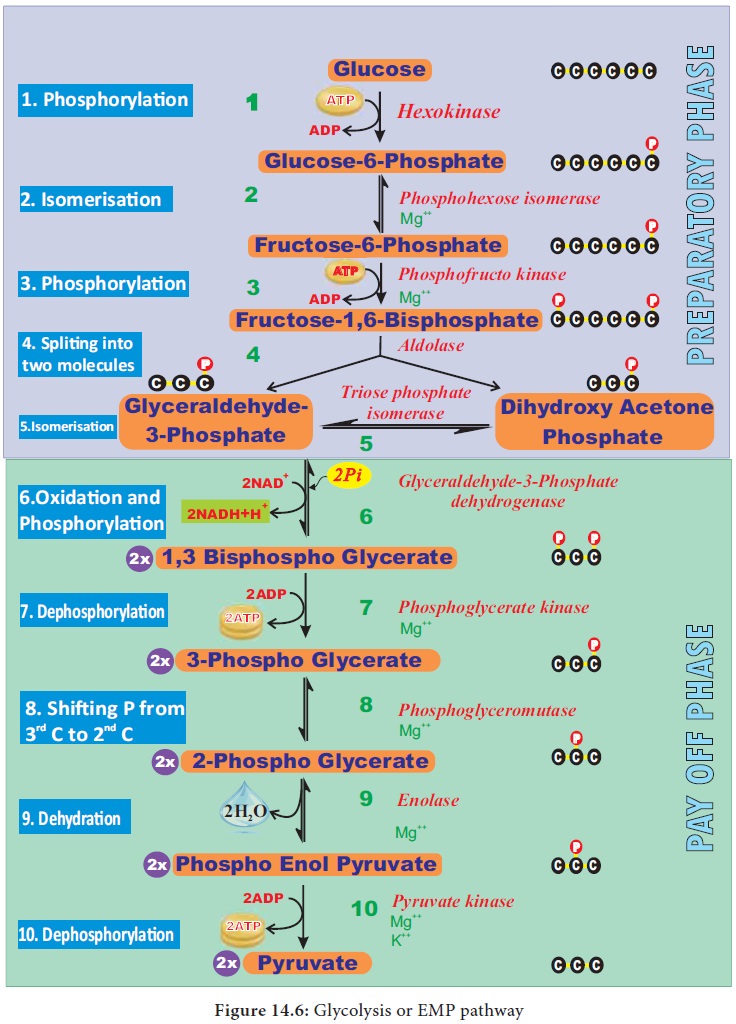
1. Preparatory phase
Glucose
enters the glycolysis from sucrose which is the end product of photosynthesis.
Glucose is phosphorylated into glucose-6-phosphate by the enzyme hexokinase,
and subsequent reactions are carried out by different enzymes (Figure 14.6). At
the end of this phase fructose-1, 6 - bisphosphate is cleaved into
glyceraldehyde-3- phosphate and dihydroxy acetone phosphate by the enzyme
aldolase. These two are isomers. Dihydroxy acetone phosphate is isomerised into
glyceraldehyde-3- phosphate by the enzyme triose phosphate isomerase, now two
molecules of glyceraldehyde 3 phosphate enter into pay off phase. During
preparatory phase two ATP molecules are consumed in step-1 and step-3 (Figure
14.6).
2. Pay off phase
Two
molecules of glyceraldehyde-3-phosphate oxidatively phosphorylated into two
molecules of 1,3 - bisphospho glycerate. During this reaction 2NAD1 is reduced
to 2NADH + H+ by glyceraldehyde-3- phosphate dehydrogenase at step
6. Further reactions are carried out by
different enzymes and at the end two molecules of pyruvate are produced. In
this phase, 2ATPs are produced at step 7 and 2 ATPs at step10 (Figure 14.6).
Direct transfer of phosphate moiety from substrate molecule to ADP and is
converted into ATP is called substrate
phosphorylation or direct
phosphorylation or trans phosphorylation . During the reaction
at step 9, 2phospho glycerate
dehydrated into Phospho enol pyruvate a water molecule is removed by the enzyme
enolase. As a result, enol group is formed within the molecule. This process is
called Enolation.
3. Energy Budget
In the
pay off phase totally 4ATP and 2NADH + H+ molecules are produced.
Since 2ATP molecules are already consumed in the preparatory phase, the net
products in glycolysis are 2ATPs and 2NADH + H+.
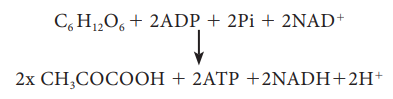
The
overall net reaction of glycolysis
2. Pyruvate Oxidation (Link reaction)
Two
molecules of pyruvate formed by glycolysis in the cytosol enters into the
mitochondrial matrix. In aerobic respiration this pyruvate with coenzyme A is
oxidatively decarboxylated into acetyl CoA by pyruvate dehydrogenase complex.
This reaction is irreversible and produces two molecules of NADH + H+
and 2CO2. It is also called transition
reaction or Link reaction. The
reaction of pyruvate oxidation is
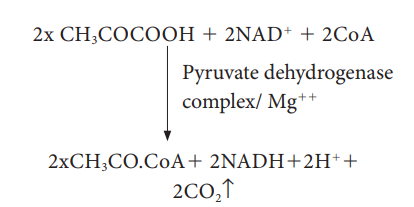
Pyruvate dehydrogenase complex consist of three distinct
enzymes, such as
1.
Pyruvate dehydrogenase
2.
Dihydrolipoyil transacetylase
3.
Dihydrolipoyil dehydrogenase and five
different coenzymes, TPP (Thymine Pyro Phosphate), NAD1, FAD, CoA and lipoate.
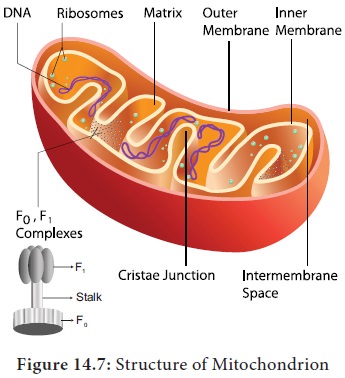
3. Krebs cycle or Citric acid cycle or TCA cycle:
Two
molecules of acetyl CoA formed from link reaction now enter into Krebs cycle.
It is named after its discoverer, German Biochemist Sir Hans Adolf Krebs (1937). The enzymes necessary for TCA cycle
are found in mitochondrial matrix except succinate dehydrogenase enzyme which
is found in mitochondrial inner membrane (Figure 14.7).
TCA cycle starts with condensation of acetyl CoA with oxaloacetate in the presence of water to yield citrate or citric acid. Therefore, it is also known as Citric Acid Cycle (CAC) or Tri Carboxylic Acid (TCA) cycle. It is followed by the action of different enzymes in cyclic manner. During the conversion of succinyl CoA to succinate by the enzyme succinyl CoA synthetase or succinate thiokinase, a molecule of ATP synthesis from substrate without entering the electron transport chain is called substrate level phosphorylation. In animals a molecule of GTP is synthesized from GDP1Pi. In a coupled reaction GTP is converted to GDP with simultaneous synthesis of ATP from ADP+Pi. In three steps (4, 5, 9) in this cycle NAD1 is reduced to NADH+ H+ and at step 7 (Figure14.8) where FAD is reduced to FADH2.
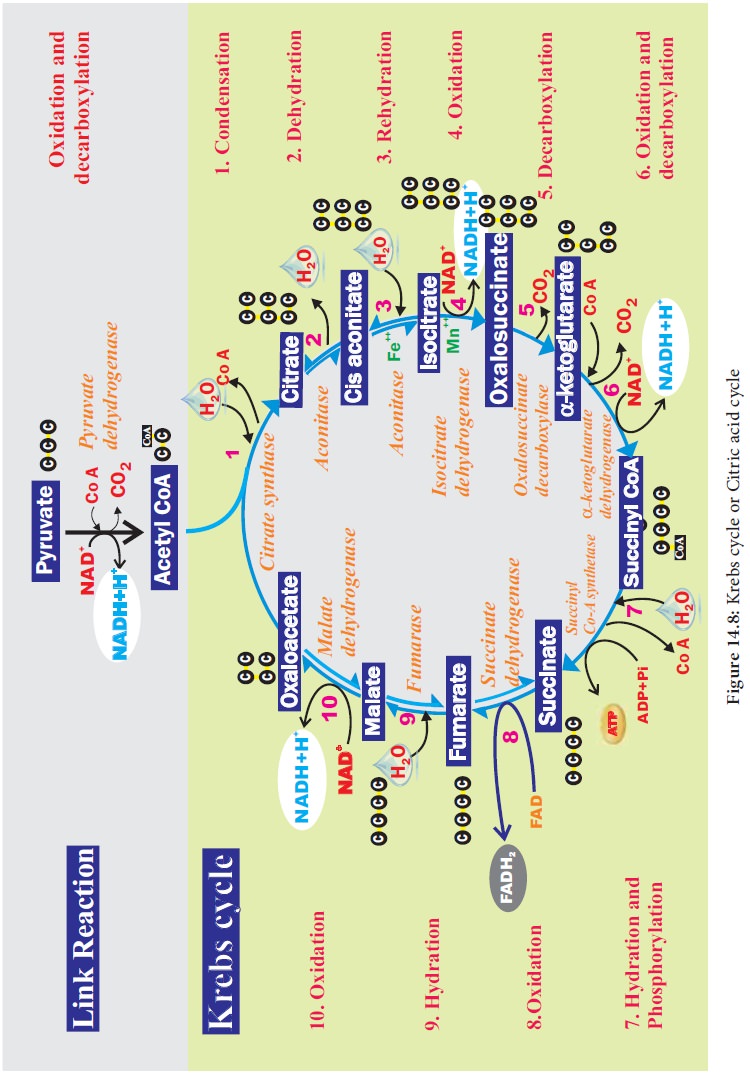
The
summary of link reaction and Krebs cycle in Mitochondria is
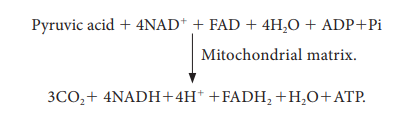
Two
molecules of pyruvic acid formed at the end of glycolysis enter into the
mitochondrial matrix. Therefore, Krebs cycle is repeated twice for every
glucose molecule where two molecules of pyruvic acid produces six molecules of
CO2, eight molecules of NADH + H+, two molecules of FADH2
and two molecules of ATP.
1. Significance of Krebs cycle:
1.
TCA cycle is to provide energy in the form of ATP
for metabolism in plants.
2.
It provides carbon skeleton or raw material for
various anabolic processes.
3.
Many intermediates of TCA cycle are further
metabolised to produce amino acids, proteins and nucleic acids.
4.
Succinyl CoA is raw material for formation of
chlorophylls, cytochrome, phytochrome and other pyrrole substances.
5.
α-ketoglutarate and oxaloacetate undergo reductive
amination and produce amino acids.
6.
It acts as metabolic sink which plays a central
role in intermediary metabolism.
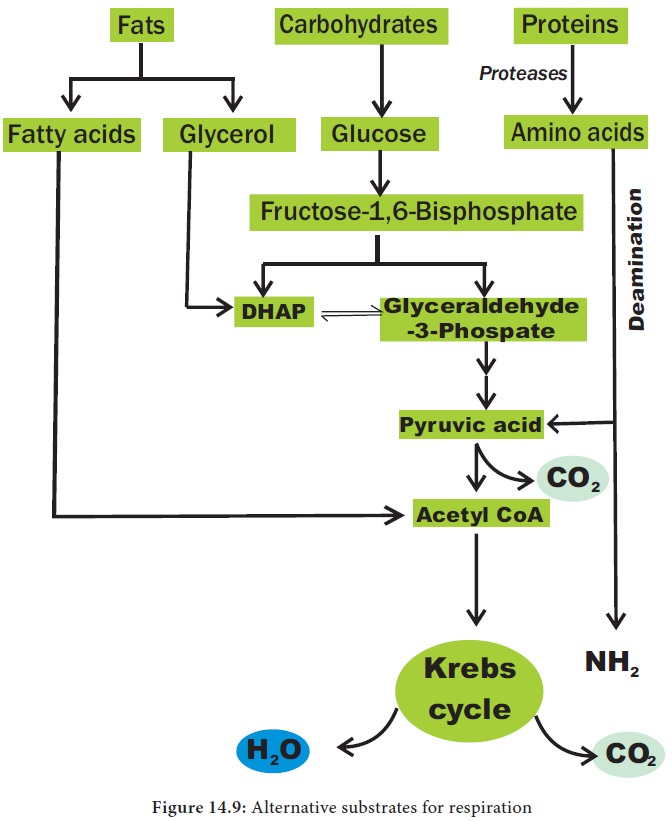
2. Amphibolic nature
Krebs
cycle is primarily a catabolic pathway, but it provides precursors for various
biosynthetic pathways there by an anabolic pathway too. Hence, it is called amphibolic pathway. It serves as a
pathway for oxidation of carbohydrates, fats and proteins. When fats are
respiratory substrate they are first broken down into glycerol and fatty acid.
Glycerol is converted into DHAP and acetyl CoA. This acetyl CoA enter into the
Krebs cycle. When proteins are the respiratory substrate they are degraded into
amino acids by proteases. The amino acids after deamination enter into the
Krebs cycle through pyruvic acid or acetyl CoA and it depends upon the
structure. So respiratory intermediates form the link between synthesis as well
as breakdown. The citric acid cycle is the final common pathway for oxidation
of fuel molecules like amino acids, fatty acids and carbohydrates. Therefore,
respiratory pathway is an amphibolic pathway (Figure 14.9).
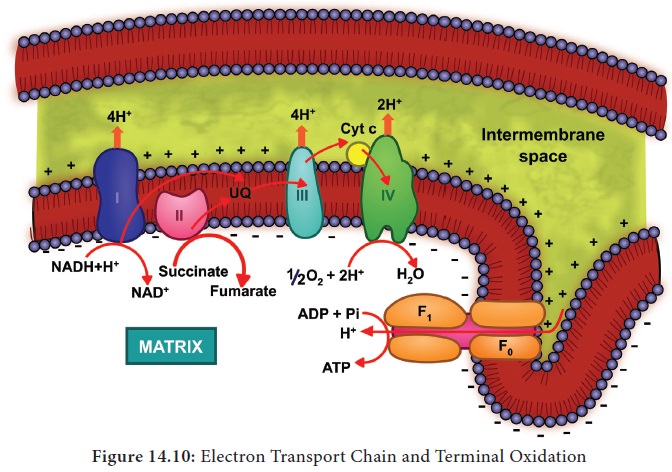
4. Electron Transport Chain (ETC) (Terminal oxidation)
During
glycolysis, link reaction and Krebs cycle the respiratory substrates are oxidised
at several steps and as a result many reduced coenzymes NADH + H+ and
FADH2 are produced. These reduced coenzymes are transported to inner
membrane of mitochondria and are converted back to their oxidised forms produce
electrons and protons. In mitochondria, the inner membrane is folded in the
form of finger projections towards the matrix called cristae. In cristae many
oxysomes (F1 particles) are present which have electron transport
carriers are present. According to Peter
Mitchell’s Chemiosmotic theory this
electron transport is coupled to ATP
synthesis. Electron and hydrogen(proton) transport takes place across four
multiprotein complexes(I-IV). They are
1.
Complex-I
(NADH dehydrogenase).
In plants, an additional NADH dehydrogenase (External) complex is present on the outer surface of inner membrane
of mitochondria which can oxidise cytosolic NADH + H+.
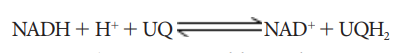
Ubiquinone
(UQ) or Coenzyme Quinone(Co Q) is a small, lipid soluble electron, proton
carrier located within the inner membrane of mitochondria.
2.
Complex-II
(Succinic dehydrogenase) It contains FAD flavoprotein is associated with
non-heme iron Sulphur (Fe-S) protein. This complex receives electrons and
protons from succinate in Krebs cycle and is converted into fumarate and passes
to ubiquinone.
Succinate
+ UQ → Fumarate + UQH2
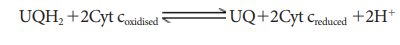
3.
Complex-III
(Cytochrome bc1 com-plex) This complex oxidises reduced
ubi-quinone (ubiquinol) and transfers the elec-trons through Cytochrome bc1
Complex (Iron Sulphur center bc1 complex) to cy-tochrome c.
Cytochrome c is a small pro-tein attached to the outer surface of inner
membrane and act as a mobile carrier to transfer electrons between complex III
to complex IV.
4. Complex IV (Cytochome c oxidase)
This
complex contains two copper centers (A and B) and cytochromes a and a3.
Complex IV is the terminal oxidase and brings about the reduction of 1/2 O2
to H2O.Two protons are needed to form a molecule of H2O
(terminal oxidation).
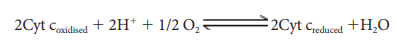
The
transfer of electrons from reduced coenzyme NADH to oxygen via complexes I to IV is coupled to the synthesis of ATP from ADP and inorganic phosphate (Pi) which is
called Oxidative phosphorylation. The F0F1-ATP
synthase (also called complex V)
consists of F0 and F1 . F1 converts ADP and Pi
to ATP and is attached to the matrix side of the inner membrane. F 0
is present in inner membrane and acts as a channel through which protons come
into matrix.
Oxidation of one molecule of NADH + H+ gives rise to 3 molecules of ATP and oxidation of one molecule FADH2 produces 2 molecules of ATP within a mitochondrion. But cytoplasmic NADH + H+ yields only two ATPs through external NADH dehydrogenase. Therefore, two reduced coenzyme (NADH + H+) molecules from glycolysis being extra mitochondrial will yield 2 x 2 = 4 ATP molecules instead of 6 ATPs (Figure 14.10) . The Mechanism of mitochondrial ATP synthesis is based on Chemiosmotic hypothesis. According to this theory electron carriers present in the inner mitochondrial membrane allow for the transfer of protons (H+). For the production of single ATP, 3 protons (H+) are needed. The terminal oxidation of external NADH bypasses the first phosphorylation site and hence only two ATP molecules are produced per external NADH oxidised through mitochondrial electron transport chain.
However, in those animal tissues in which malate
shuttle mechanism is present, the oxidation of external NADH will yield almost
3 ATP molecules.
Abnormal rise in respiratory rate of ripening in fruits is
called Climacteric. Examples are
apple, banana, mango, papaya, pear.
Complete
oxidation of a glucose molecule in aerobic respiration results in the net gain
of 36 ATP molecules in plants as
shown in table 14.2. Since huge amount of energy is generated in mitochondria
in the form of ATP molecules they are called ‘power house of the cell’.
In the case of aerobic prokaryotes due to lack of mitochondria each molecule of
glucose produces 38 ATP molecules.
Recent view
When the
cost of transport of ATPs from matrix into the cytosol is considered, the
number will be 2.5 ATPs for each NADH + H+ and 1.5 ATPs for each
FADH2 oxidised during electron transport system. Therefore, in
plant cells net yield of 30 ATP molecules for complete aerobic oxidation of one
molecule of glucose. But in those animal cells (showing malate shuttle
mechanism) net yield will be 32 ATP molecules.
Electron transport chain inhibitors
1. 2,4 DNP (Dinitrophenol) - It
prevents synthesis of ATP from ADP,
as it directs electrons from Co Q to O2
2. Cyanide - It prevents flow of electrons from Cytochrome a3 to O2
3. Rotenone - It prevents flow of electrons from NADH + H+/FADH2 to Co
Q
4. Oligomycin – It inhibits oxidative phosphorylation
Related Topics