Chapter: Aquaculture Principles and Practices: Nutrition and Feeds
Proteins - Aquaculture: Energy requirements and sources
Proteins
Dietary protein is the main source of nitrogen and essential amino acids in animals. It is also the most expensive source of energy in artificial diets. In nature, carnivorous fish consume foods which are about 50 per cent protein. They have a very efficient system for excretion of waste nitrogen from protein, which is catabolized for energy. Therefore high-protein diets are not harmful but, being expensive, it is necessary to keep the proportion of protein down to optimum levels necessary for good growth and feed conversion. Protein has a metabolizable energy value of about 4.5 kcal/g in fish, which is higher than that of mammals and birds.
Gross protein requirements of a number of cold-water and warm-water finfish have been determined (Table 7.1). The requirements are highest in the initial feeding of fry and decrease as fish size increases. For maximum growth, young fish require between 40 and 60 per cent of their diet as proteins, which is much higher
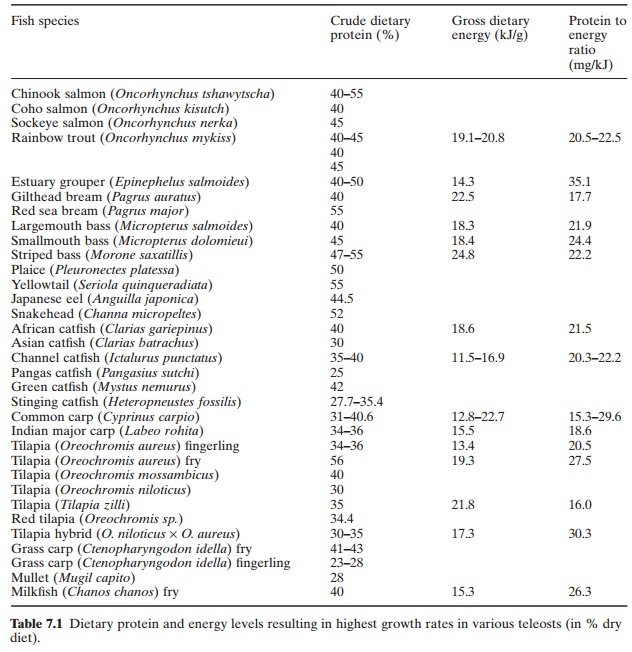
than the requirements of terrestrial animals. However, most of the wet weight gain in lean fish is in the form of muscle tissue, unlike in terrestrial animals where there is considerable deposition of both fat and protein. Salmonids continue to need, from young to adult stages, higher levels of 40–60 per cent protein in their diets. But other species like the milkfish (Chanos chanos) appear to make a rapid tran sition to a diet of algae, containing 10–20 per cent protein in its natural environment. The protein component of this material is digested and the amino acids absorbed, while most of the undigestible cellulose is excreted. The dietary protein level resulting in the highest growth rate in various farmed shrimp species ranges from 28–57 per cent, the highest requirement being 40–58 per cent and 40–51 per cent for Penaeus japonicus and P. aztecus respectively and the lowest 28–32 per cent for P.setiferus. The optimal growth ranges for P. inducus, P. merguiensis and P. monodon werefound to be with 30–40 per cent, 32–42 per cent and 34–46 per cent diets respectively. The corresponding range for the freshwater prawn,
Macrobrachium rosenbergii, is 35–40 per cent(Tacon, 1990).
Protein requirements are influenced by water temperature, body size, stocking density, oxygen levels and the presence of toxins. As water temperature declines, the body temperature of fish also declines and consequently the metabolic rate is reduced. The most favourable temperature for a given species is the one at which the difference between maintenance requirement and voluntary food intake is greatest and at which optimum efficiency of growth occurs (Smith, 1980). Chinook salmon (Oncorhynchus tshawytscha) need food containing 40 per cent protein in water temperatures of about 8°C for optimum growth, whereas in temperatures of about 14°C the same fish will need food containing 55 per cent protein (De Long et al., 1958). At lower temperatures, foods containing more than 40 per cent protein produce stress due to an excess of ammonia released from gills. Channel catfish (Ictalurus punctatus) show optimum growth at 20°C on a 35 per cent protein diet, whereas at 25°C they need a 40 per cent protein diet to achieve optimum growth (Dupree and Sneed, 1966). However, work by Slinger et al. (1977) and Cho and Slinger (1978), does not confirm the results relating to temperature effects. The greater absolute need for protein at higher temperatures might be satisfied through increased consumption of the lower protein diets Readily digested high-protein materials have higher metabolizable energy (ME) values for fish than other mono-gastric animals. Similarly, protein has more net energy for fish than it has for mammals or birds. Smith et al. (1978) showed that less than 5 per cent of the ME is lost as heat increment in fish. Fish are among the most efficient of all animals in converting feed energy into high quality protein.
Available information does not seem to support the general view that omnivorous and herbivorous aquaculture species require lessprotein in their diets. For example, the juveniles of herbivorous grass carp (Ctenopharyngodonidella) require levels of protein similar tosalmon and trout. Phytoplankton and zoo-plankton contain high percentages of protein (40–60 per cent) and there is reason to believe that the protein requirements of plankton-feeding species are also similarly high. The real difference between species of different feeding habits would appear to be in the ability to digest carbohydrates. Most of the carnivores, like trout and yellowtail, have a limited ability to digest complex carbohydrates.
Juveniles and adults of most cultured crustaceans have protein requirements in the range of 30–50 per cent of their dry diet weight. Like fish, they also require much higher levels of protein than terrestrial animals. However, there are differences in their nitrogen metabolism. One major difference is based on the habit of moulting. The crustacean exoskeleton consists of a mineral-organic matrix. Chitin, one of the primary compounds, is composed of glucosamine units (an amine group and glucose). Growth occurs when the old exoskeleton is partly resorbed, then shed and a new one grown in its place. Prior to moulting, they produce high levels of ammonia, indicating the resorption of the old exoskeleton. Even when the exuvia is eaten, substantial losses of nitrogen occur. Nitrogen balance can be used to evaluate the amino acid and nitrogen requirements. Moulting (ecdysis) affects the animal’s nitrogen balance.
Unlike terrestrial vertebrates, crustaceans appear to have a limited ability to store protein (Maynard and Loosi, 1969). Recent studies show that both carbohydrates and lipids can be used to spare dietary protein in crustaceans. There are wide variations in the protein requirements of shrimp species Little work has been done on defining the nutritional requirements of molluscs, probably because of the successful use of algae as food and the lack of successful microencapsulated diets. However, as pointed out earlier, the protein content of phytoplankton species is generally high (above 40 per cent), although it varies with environmental conditions.
Studies by Langton et al. (1977) on the relationship between protein consumed andnitrogen retained by the clam Tapes japonicashowed that they are directly correlated. However, the efficiency dropped from 48 to 36 per cent when the nitrogen intake doubled from approximately 50 to 100 mg. Its nitrogen retention efficiency thus appears to be similar to that of other aquatic animals.
Related Topics