Chapter: Human Nervous System and Sensory Organs : Functional Systems
Olfactory Organ

Olfactory Organ
In
humans, the olfactory epithelium
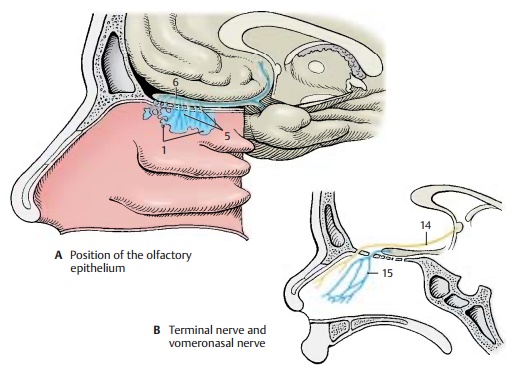
occu-pies a small region in both nasal cavities (ol-factory region) (A1)
at the upper margin ofthe superior nasal concha and on the opposite surface of
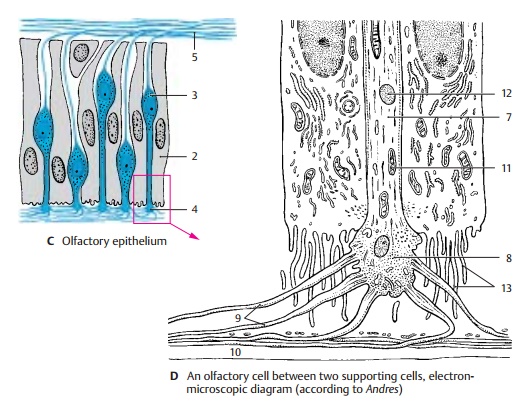
the nasal septum. The mul-tilayered sensory epithelium is composed of supporting cells (C2) and receptor cells (C3)that are characterized by pale,
deep-lying cell nuclei. The olfactory region also con-tains small mucous
glands, the olfactoryglands (Bowman’s glands). Their secretioncovers
the olfactory mucosa as a thin film.
The
apical part of the sensory cell tapers into a thin shaft that slightly extends
be-yond the surface of the epithelium. This knoblike olfactory vesicle (C4)
is occupied by a number of olfactory
hairs. At the basal end, the ovoid cell body forms a fine process that,
together with several other processes, is en-veloped by Schwann cells. The
bundled processes (fila olfactoria)
represent the ol-factory nerves (AC5) and extend through theopenings of
the cribriform plate to the ol-factory
bulb (A6). In the olfactorybulb,
the processes terminate in the ol-factory
glomeruli, where they form synapticcontacts with the dendrites of mitral cells. The epithelial sensory
cells are bipolar neu-rons; the short dendrite represents the re-ceptor part
and the long axon runs as centripetal fiber to the olfactory bulb.
Electron
micrographs of olfactory cells (in the cat) show that the apical shaft of the
cell (D7) terminates with a knob (D8) from which numerous long olfactory cilia (D9) ex-tend. The terminal parts of the sensory cilia lie in the
mucous layer (D10) that seals the
entire surface of the olfactory epithelium against the air space. Shaft and
knob contain microtubules, numerous mitochondria (D11), and some lysosomes (D12).
The knob extends above the surface of the supporting cells, which exhibit a
dense border of micro-villi (D13).
How the
sensory cells receive the different odors is currently the topic of intensive
re-search. The odoriferous substances must be water-soluble in order to
dissolve in thesuperficial mucous layer and reach the sensory cilia where they
bind to specific membrane receptors. In sufficiently high concentrations, they
induce depolarization of the membrane, which is conducted as an action
potential in the cell’s axon. As in the case of taste, it is assumed that there
are a few basic qualities of odor and that one sensory cell registers only one
particular ba-sic quality via specific receptors. Since sub-stances belonging
to a group of odors have roughly the same molecular size, it seems possible
that the membrane of an olfactory cilium reacts to only one particular
molecu-lar size. Indeed, recent studies suggest that each sensory cell
expresses only one recep-tor type. In the mouse, the sensory cells of one
receptor type project to only two of the 1800 glomeruli in the olfactory bulb
(central olfactory pathway,).
Apart
from the olfactory nerves, two other paired nerves run from the nasal cavity to
the brain, namely, the terminal nerve and the vomeronasal nerve. The terminal nerve (B14) consists of a bundle of delicate nerve fibers that extends
from the nasal septum through the cribriform plate to the terminal lamina and
enters below the anterior com-missure into the brain. The bundle contains
numerous neurons and is regarded as an au-tonomic nerve. The vomeronasal nerve (B15), which runs from the vomeronasal
organ (Ja-cobson’s organ) to the
accessory olfactorybulb, is well developed in lower vertebrates but can be
demonstrated in humans only during embryonic development. In reptiles, the
vomeronasal organ is a sensory epithelium in a pocket of the mucosa of the
nasal septum; it is thought to play an impor-tant role in tracking down prey.
Related Topics