Chapter: Human Nervous System and Sensory Organs : Functional Systems
Muscle Spindle - Motor Systems
Muscle Spindle
A muscle
spindle, or neuromuscular spindle, consists of 5 – 10 thin striated muscle
fibers (intrafusal muscle fibers) (A1) which are surrounded by a
fluid-filled connective-tissue capsule (A2).
The fibers of the up to 10 mm long spindles are arranged in parallel with the
other fibers of the muscle (extrafusal
muscle fibers) and attach either to the tendons of the muscle or to the
connective-tissue poles of the capsule. As the intrafusal fibers lie in the
same longi-tudinal orientation as the extrafusal fibers, stretching and
shortening of the muscle af-fects them in the same way. The number of spindles
within individual muscles is quite variable. Muscles participating in delicate
and precise movements (finger muscles) possess a large number of spindles,
whereas muscles for simple movements (trunk muscles) contain far fewer
spindles.
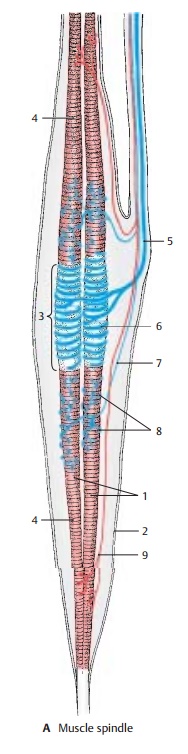
The
central equatorial part (A3) of an
intra-fusal fiber contains several cell nuclei but no myofibrils; this part of
the spindle is not contractile. Only the two segments (A4) that contain striated myofibrils are contractile. A thick
sensory nerve fiber (A5) terminates
at the central part; its terminal branches wind around the muscle fibers like
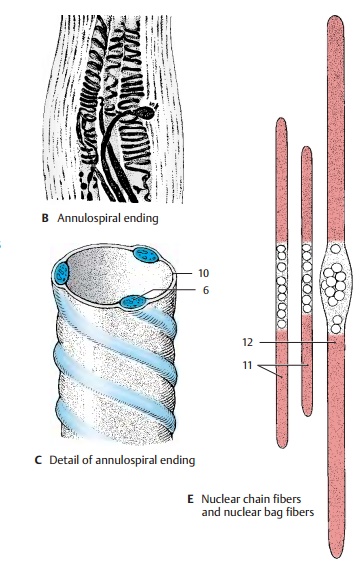
spirals and form the annulospiral
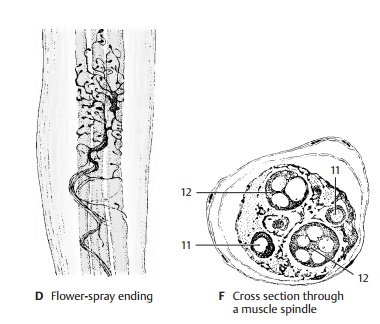
endings (AC6; B). A delicate sensory fiber (A7) attaches in an umbelliform fashion
(flower-spray ending) (A8, D) at one side, or at both sides, of the annulospiral ending.
Both
contractile polar segments are inner-vated by thin fusimotor fibers (γ-fibers) (A9). Their small motor end plates have only poorly developed
subneural clefts; like the extrafusal muscle fibers, they are epilem-mal. The sensory annulospiral
endings liebelow the basement membrane of the muscle fiber (C10) and, hence, are hypolem-mal. Theγ-fibers stem from small
motorneurons in the anterior horn (γ-mo-toneurons); impulses from these neurons
cause contraction of the polar segments of the intrafusal fiber. This results
in stretching of the equatorial segment and does not only stimulate the
annulospiral ending but alters the sensitivity of the spindle as well.
The
muscle spindle is a stretch receptor,
which is stimulated when the muscle is stretched but becomes inactive when the
muscle is contracted. Upon stretching the muscle the impulse frequency
increases with the change in muscle length. This way, the spindles transmit
information on the prevailing length of the muscle. The im-pulses are
transmitted not only via the spinocerebellar tracts to the cerebellum but also
via reflex collaterals directly to the large anterior horn cells
(α-motoneurons). Stimulation of the latter neurons during sudden stretching
results in immediate muscle contraction (stretch
reflex).
The
muscle spindle contains two different types of intrafusal fibers: the nuclear chainfibers (EF11) and the nuclear bag fibers (EF12).Both
types of fiber are innervated by an-nulospiral endings. Flower-spray endings
are predominantly found at nuclear chain fibers. The thicker nuclear bag fibers
re-spond to the ongoing stretching of the muscle, whereas the continuous state
of muscle stretching is registered by the thin-ner nuclear chain fibers. Thus,
muscle spindles transmit to the cerebellum not only information on the length
of the muscle but also on the speed of muscle stretching.
Apart
from tendon organs and muscle spindles, there are sensory end organs in joint
capsules and ligaments (tension
recep-tors) that constantly send information onmovement and posture of
trunk and limbs to the cerebellum (anterior and posterior spinocerebellar
tracts).
Related Topics