Chapter: Introduction to Human Nutrition: The Vitamins
Absorption and metabolism of vitamin B6
Absorption and metabolism
The phosphorylated vitamers are dephosphorylated by membrane-bound alkaline phosphatase in the intestinal mucosa; pyridoxal, pyridoxamine, and pyri-doxine are all absorbed rapidly by passive diffusion. Intestinal mucosal cells have pyridoxine kinase and pyridoxine phosphate oxidase (Figure 8.11), so that there is net accumulation of pyridoxal phosphate by metabolic trapping. Much of the ingested pyridoxine is released into the portal circulation as pyridoxal, after dephosphorylation at the serosal surface. Unlike other B vitamins, there seems to be no limit on the amount of vitamin B6 that is absorbed.
Most of the absorbed vitamin is taken up by the liver by passive diffusion, followed by metabolic trap-ping as phosphate esters, which do not cross cell membranes, then oxidation to pyridoxal phosphate. The liver exports both pyridoxal phosphate (bound to albumin) and pyridoxal (which binds to both albumin and hemoglobin). Free pyridoxal remaining in the liver is rapidly oxidized to 4-pyridoxic acid, which is the main excretory product.
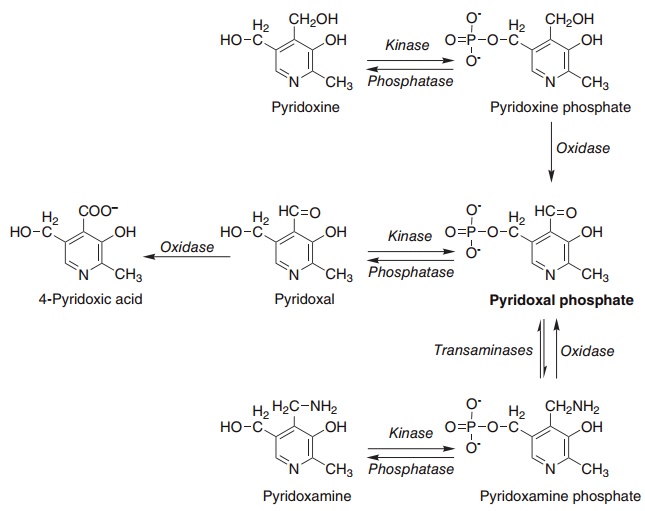
Figure 8.11 Interconversion of the vitamin B6 vitamers. Pyridoxal kinase (EC 2.7.1.38), pyridoxine phosphate oxidase (EC 1.1.1.65), pyridoxamine phosphate oxidase (EC 1.4.3.5).
Extrahepatic tissues take up pyridoxal and pyri-doxal phosphate from the plasma. The phosphate is hydrolyzed to pyridoxal, which can cross cell mem-branes, by extracellular alkaline phosphatase, then trapped intracellularly by phosphorylation. Tissue concentrations of pyridoxal phosphate are controlled by the balance between phosphorylation and dephosphorylation.
Some 80% of the body’s total vitamin B6 is pyri-doxal phosphate in muscle, mostly associated with glycogen phosphorylase. This does not function as a reserve of the vitamin and is not released from muscle in times of deficiency; it is released into the circula-tion (as pyridoxal) in starvation, when glycogen reserves are exhausted and there is less requirement for phosphorylase activity. Under these conditions it is available for redistribution to other tissues, and especially the liver and kidneys, to meet the increased need for transamination of amino acids to provide substrates for gluconeogenesis.
Related Topics