Chapter: Genetics and Molecular Biology: Genetic Engineering and Recombinant DNA
Vectors for Higher Cells
Vectors for Higher Cells
Cloning DNA in higher cells poses the same problems
as cloning in bacteria. The vectors must permit simple purification of sizable
quan-tities of DNA, must permit selection of transformed cells, and must have
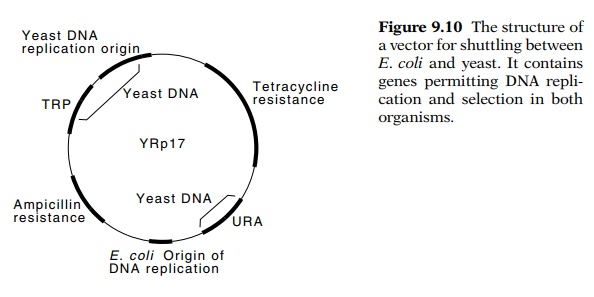
space for inserted DNA. Shuttle vectors, which have been extensively used for
cloning in yeast, are a neat solution to these requirements. In addition to containing
the normal bacterial cloning-plasmid elements, they contain a yeast replicon
and a genetic marker selectable in yeast (Fig. 9.10). As a result, large
quantities of the vectors can be obtained by growth in E. coli and then
transformed into yeast. The ability to shuttle between bacteria and yeast saves
much time and expense in genetic engineering experiments.
Two types of yeast replication origins can be used
in yeast shuttle vectors. One is a yeast chromosomal DNA replication origin,
also known as an ARS element. The other is the origin from the 2 µ circles. These are plasmid-like elements with unknown function that are
found in yeast. They are somewhat more stable than the ARS vectors. Nutritional
markers such as uracil, histidine, leucine, and tryptophan biosynthesis have
been used as selectable genes in the appropriately auxotrophic yeast.
Viruses form a basis for many vectors useful in
higher plant and animal cells. For example, one of the simplest vectors for
mammalian cells is the simian virus SV4O. It permits many of the same cloning
operations as phage lambda.
The terminology used with mammalian cells can be
confusing. “Transformation” can mean that cells have received a plasmid. It can
also mean that the cells have lost their contact inhibition. In this state they
continue growing past the confluent cell monolayer stage at which
Figure
9.10 The structure ofa vector for
shuttling between E. coli and yeast. It containsgenes
permitting DNA repli-cation and selection in both organisms.
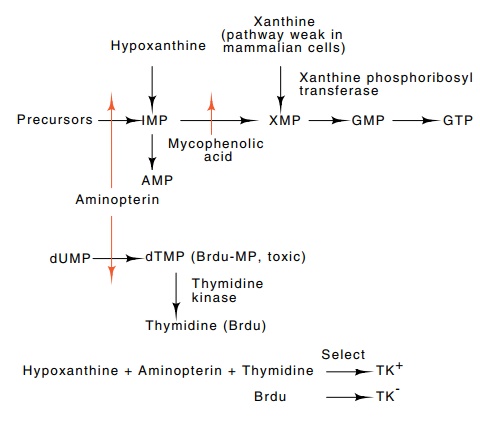
Figure
9.11 The metabolic pathways involved
with some selectable genes inmammalian cells. IMP-inosine monophosphate,
XMP-xanthine monophos-phate, GMP-guanosine monophosphate, dUMP-deoxyuridine
monophosphate, dTMP-deoxythymidine monophosphate, Brdu-bromodeoxyuridine,
Brdu-MP, bromodeoxyuridine monophosphate. Aminopterin blocks tetrahydrofolate
re-ductase, which is required for synthesis of IMP and dTMP, and mycophenolic
acid blocks synthesis of XMP.
normal
mammalian cells cease growth. Transformation to the uninhibi-ted growing state
can result from infection by a tumor-causing virus like SV4O or it can be a
result of mutation of the genome. Although loss of contact inhibition could be
useful in identifying cells that have incorpo-rated the SV4O DNA or SV4O
hybrids, this property is of limited use. Other selectable genetic markers
suitable for mammalian cells are required.
One
useful gene for selections in mammalian cells has been the thymidine kinase gene
because TK+ cells can be selected by growing them in medium
containing hypoxanthine, aminopterin, and thymid-ine. Conversely, TK-
cells can be selected by growing them in medium containing bromodeoxyuridine
(Fig. 9.11). Furthermore, virologists had previously discovered that the herpes
simplex virus codes for its own thymidine kinase. Therefore the viral genome
can be used as a concentrated source of the gene in an expressible form for
initial cloning experiments.
Although
the thymidine kinase gene has been useful in selecting cells that have taken up
foreign DNA, a selectable gene that does not require the prior isolation of a
thymidine kinase negative mutant in each cell line would also be valuable. The E. coli
enzyme xanthine-guanine
phosphoribosyl
transferase gene appears to meet these requirements. The protein product of the
gene functions in mammalian cells and permits selective growth of nonmutant
cells that contain the enzyme (Fig. 9.11). The required growth medium contains
xanthine, hypoxan-thine, aminopterin, and mycophenolic acid. Other dominant
genes useful for the selection of transformed cells are mutant dihydrofolate
reductase that is resistant to methotrexate, a potent inhibitor of the
wild-type enzyme, and kanamycin-neomycin phosphotransferase. The latter is an
enzyme derived from a bacterial transposon and confers resistance to bacteria,
yeast, plant, and mammalian cells to a compound called G418 . Of course, for
proper expression in the higher cells the gene must be connected to an appropriate
transcription unit and must contain the required translation initiation and
polyadenylation signals.
Related Topics