Chapter: Biochemistry: Viruses, Cancer, and Immunology
T-Cell Functions
What do T cells and B cells do?
T-Cell Functions
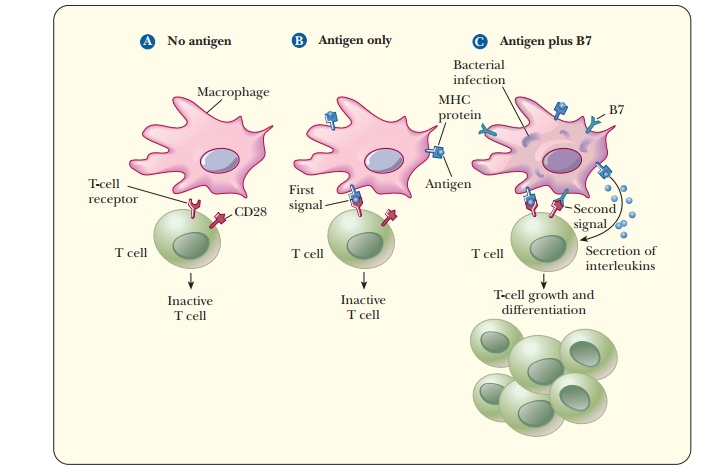
T cells can have a number of functions. As T cells differentiate, each becomes specialized for one of the possible functions. The first of these possibilities, that of the killer T cells, involves T-cell receptors (TCRs) on their surfaces that recognize and bind to antigens, the foreign substances that trigger the immune response. The antigens are presented to the T cell by antigen-presenting cells (APCs), such as macrophages and dendritic cells. The APCs ingest and process antigens, and then present them to the T cells. The processed antigen takes the form of a short peptide bound to an MHC I protein on the surface of the APC. Figure 14.14 shows how this works for macrophages.
The
macrophage also presents another molecule, a protein of a family known as B7,
which binds to another T-cell surface protein called CD28; the exact nature of
this B7 protein is a subject of active research. The combination of the two
signals leads to T-cell growth and differentiation, producing killer T cells.
Proliferation of killer T cells is also triggered when macrophages bound to T
cells produce small proteins called interleukins.
The T cells make an interleukin-receptor protein as long as they are bound to
the macrophage but do not do so when they are no longer bound. Interleukins are
part of a class of substances called cytokines. When we discussed innate
immunity, we saw that this term refers to soluble protein factors produced by
one cell that specifically affect another cell. In this way, T cells do not
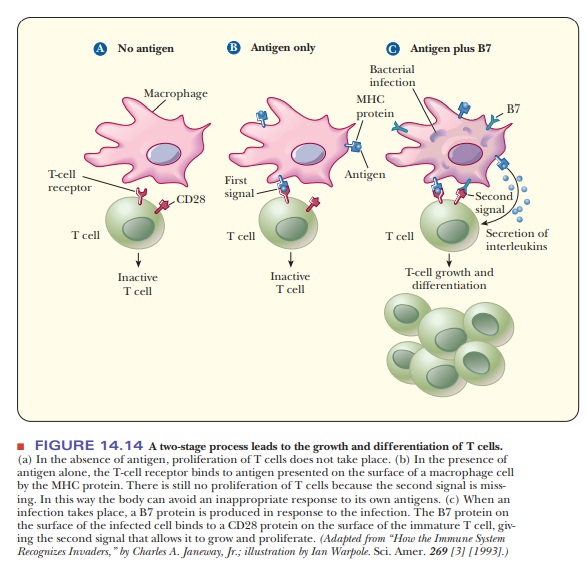
proliferate in uncontrolled fashion. A killer T cell also has another membrane
protein called CD8, which helps it dock to the MHC of the antigen-presenting
cell, as shown in Figure 14.15. In fact, the CD8 protein is such a
distinguishing characteristic that many researchers use the term CD8cells instead of killer T cells.
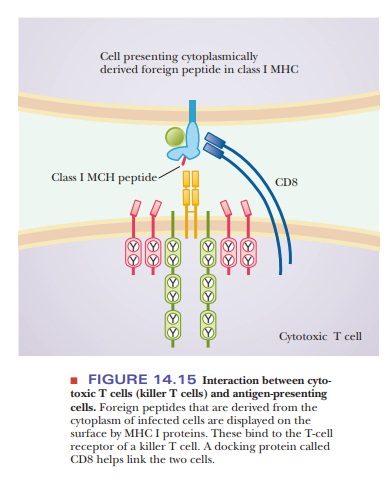
T cells that bind to a given antigen and only to that antigen grow when these conditions are fulfilled. Note the specificity of which the immune system is capable. Many substances, including ones that do not exist in nature, can be antigens. The remarkable adaptability of the immune system in dealing with so many possible challenges is another of its main features. The process by which only the cells that respond to a given antigen grow in preference to other T cells is called clonal selection (Figure 14.16).
The immune system can thus be versatile in its
responses to the challenges it meets. This clonal selection is the basis of the
definition of acquired immunity. Most of the response of T cells stems from the
rapid proliferation of cells once they are selected—the organism, in essence,
acquiring these cells only when necessary. However, there must be at least one
cell with the proper TCR to recognize the antigen and to bind to it. These
receptors are not generated because there is a need; rather, they are generated
randomly when stem cells differentiate into T cells. Fortunately, the diversity
of T-cell receptors is so great that there are millions of TCR specificities.
The division of T cells during the peak of the immune response is very rapid, often reaching three or four divisions per day. This would lead to more than a thousandfold increase in the number of selected T cells in a few days.
As their name implies, killer T cells destroy antigen-infected
cells. They do so by binding to them and by releasing a protein that perforates
the plasma membranes of the infected cells. This aspect of the immune system is
particu-larly effective in preventing the spread of viral infection by killing
virus-infected host cells. In a situation such as this, the antigen can be
considered all or part of the coat protein of the virus. When the infection
subsides, some memory cells remain, conferring immunity against later attacks
from the same virus.
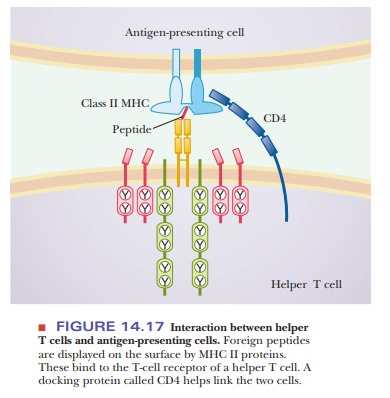
T cells play another role in the immune system. Another class of T
cells develops receptors for a different group of antigen-presenting MHC
proteins, in this case MHC II. These become helper T cells, which develop in much the same way as killer T
cells. Helper T cells are also referred to as CD4 cells, because of the presence of that unique membrane protein.
CD4 helps the cell dock to the MHC of the antigen-presenting cell, as shown in
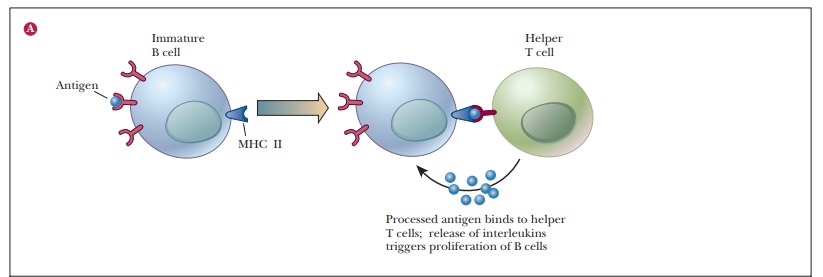
Figure 14.17. The func-tion of helper T cells is primarily to aid in the
stimulation of B cells. Maturing B cells display the MHC II protein, with
processed antigen, on their surfaces. Note particularly that the MHC proteins
play a key role in the immune system. This property has led to a considerable
amount of research to determine their structure, including determination by
X-ray crystallography. The MHC II of the B cells is the binding site for helper
T cells. The binding of helper T cells to B cells releases interleukins (IL-2
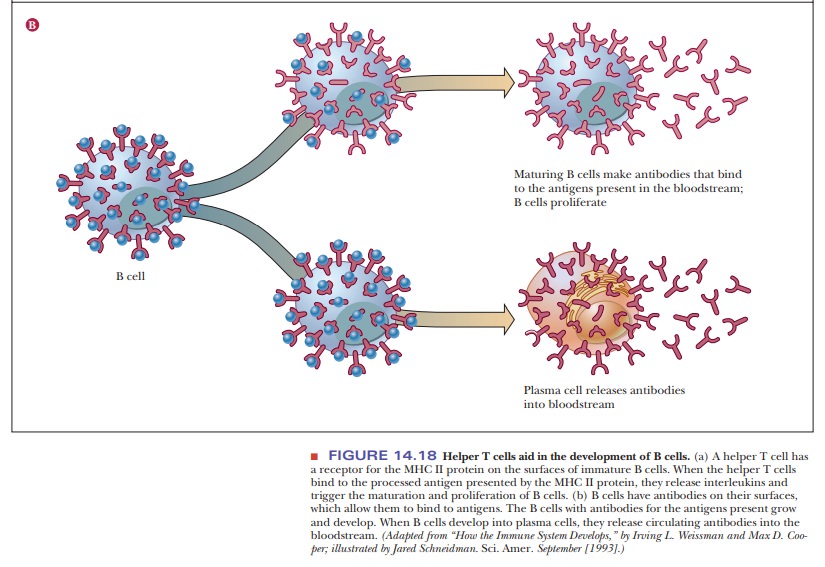
and IL-4) and triggers the development of B cells into plasma cells (Figure
14.18). Both B cells and plasma cells pro-duce antibodies (also known as immunoglobulins),
the proteins that will occupy most of our time as we discuss the molecular
aspects of the immune response. B cells display antibodies on their surfaces in
addition to the MHC II proteins. The antibodies recognize and bind to antigens.
This property allows B cells to absorb antigens for processing. Plasma cells
release circulating antibodies into the bloodstream, where they bind to antigen,
marking it for destruction by the immune system. Helper T cells also help
stimulate killer T cells and antigen-presenting cells via release of
interleukins.
Related Topics