Chapter: Human Nervous System and Sensory Organs : Diencephalon
Structure of Diencephalon
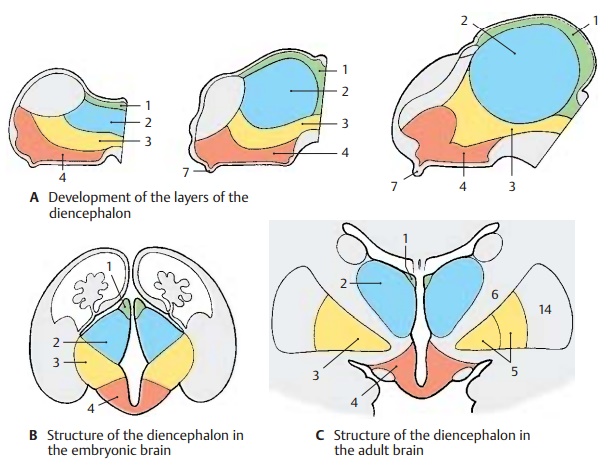
Structure
Subdivision (A – C)
The diencephalon is subdivided into four layers lying on top of each other:
! The epithalamus (A – C1)
! The dorsal thalamus (A – C2)
! The subthalamus (A – C3)
! The hypothalamus (A – C4)
The simple arrangement of these layers is still clearly visible in the embryonic brain. However, it changes considerably during development owing to differences in re-gional growth. In particular, the extraordi-nary increase in mass of the dorsal thalamus and the expansion of the hypothalamus in the region of the tuber cinereum determine the structure of the diencephalon.
The epithalamus consists of the habenulae, a relay station for pathways be-tween the olfactory centers and the brain stem, and of the pineal gland (epiphysis cerebri). Owing to the increasing size of the thalamus, the dorsally situated epithalamus (B1) becomes medially transposed and ap-pears only as an appendage of the dorsal thalamus (C1).
The dorsal thalamus is the terminal station of sensory pathways (cutaneous sensibility; taste; visual, acoustic, and vesti-bular pathways). It is connected to the cere-bral cortex by efferent and afferent fiber systems.
The subthalamus is the continua-tion of the midbrain tegmentum. It contains nuclei of the extrapyramidal motor system (zona incerta, subthalamic nucleus, globus pallidus) and may be regarded as the motor zone of the diencephalon.
The globus pallidus, or pallidum (CD5), is a derivative of the diencephalon. It becomes separated from the other gray regions of the diencephalon as a result of the ingrowing fiber masses of the internal capsule (CD6) during development and finally becomes displaced into the telencephalon. Only a small medial rest of the pallidum remains within the unit of the diencephalon; this isthe entopeduncular nucleus. As a constitu-ent of the extrapyramidal system, the globus pallidus should logically be regarded as part of the subthalamus.
The hypothalamus is derived from the lowest layer and forms the floor of the diencephalon from which the neurohy-pophysis (A7) protrudes. It is the highest regulatory center of the autonomous nervous system.
Frontal Section at the Level of the Optic Chasm (D)
A section through the anterior wall of the third ventricle shows parts of the dien-cephalon and telencephalon. Ventrally lies the fiber plate of the decussation of the optic nerve, the optic chasm (D8). A rostral excavation of the third ventricle, the preop-tic recess (D9), is seen above it. The globuspallidus (CD5) appears laterally to the inter-nal capsule. All other structures belong to the telencephalon: the two lateral ven-tricles (D10) and the septum pellucidum (D11) enclosing the cave of the septum pel-lucidum (D12), the caudate nucleus (D13),the putamen (CD14), and at the base, the ol-factory area (D15) (anterior perforated sub-stance). The corpus callosum (D16) and the anterior commissure (D17) connect the twohemispheres. Other fiber systems shown in the section are the fornix (D18) and the lateral olfactory stria (D19).
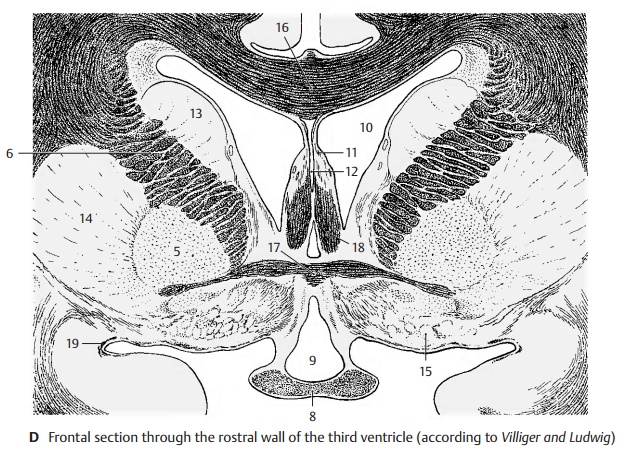
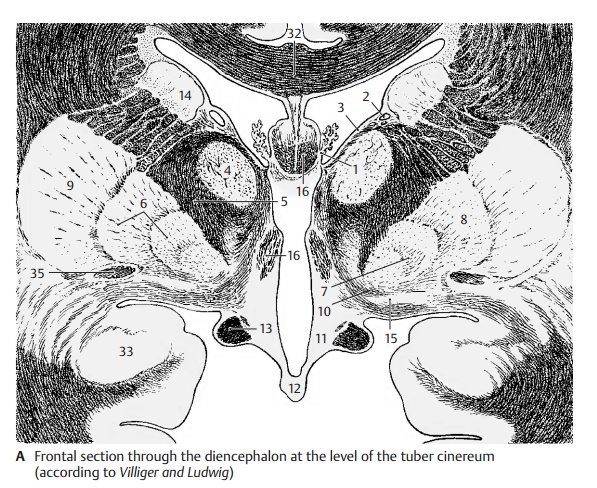
Frontal Section Through the Tuber Cinereum (A)
The plane of section lies just behind the in- terventricular foramen (foramen of Monro). Lateral ventricle and third ventricle are sep- arated by the thin base of the choroid plexus (A1). From here to the thalamostriate vein (AB2) extends the lamina affixa (AB3). It covers the dorsal surface of the thalamus (A4), of which only the anterior nuclei are visible. Ventrolaterally to it and separated by the internal capsule (AB5) lies the globus pallidus (AB6), which is divided into two parts, the internal (A7) and the external seg- ment of the pallidum (A8). It stands out against the adjacent putamen (AB9) because of its higher myelin content. At the basal margin and at the tip of the pallidum there exit the lenticular fasciculus (Forel’s fieldH2) and the lenticular ansa (A10). The latter forms an arch in dorsal direction around the medial tip of the pallidum. The ventral part of the diencephalon is occupied by the hy- pothalamus (tuber cinereum [A11] and in- fundibulum [A12]), which appears markedly poor in myelin, in contrast to the heavily myelinated optic tract (AB13). The diencephalon is enclosed on both sides by the telencephalon, but without a clearly visible boundary. The closest nuclei of the telencephalon are the putamen (AB9) and the caudate nucleus (AB14). Anterior to theglobus pallidus lies a nucleus belonging to the telencephalon, the basal nucleus (Mey-nert’s nucleus) (A15). It receives fibers fromthe midbrain tegmentum. Its large choliner-gic neurons project diffusely into the entire neocortex. The fornix (AB16) is seen twice because of its arched course.
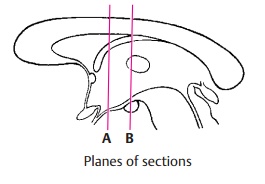
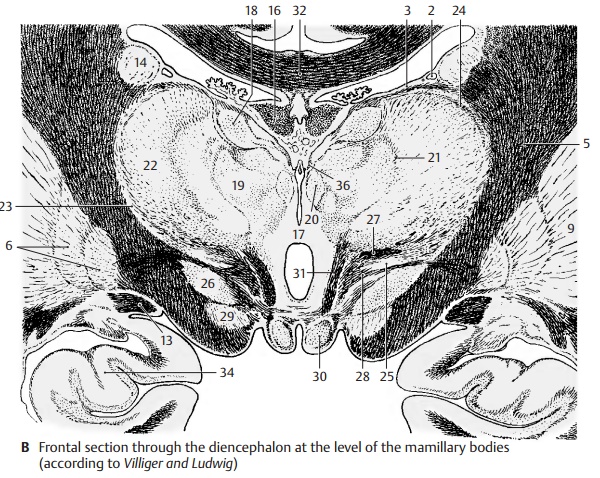
Frontal Section at the Level of the Mamillary Bodies (B)
The section shows both thalami; their in-crease in volume has lead to secondary fu-sion in the median line, resulting in the in-terthalamic adhesion (B17). Myelinated fiberlamellae, the medullary layers of thelies the anterior nuclear group (B18), ven- trally to it the medial nuclear group (B19), which borders medially on several small paraventricular nuclei (B20) and is laterally separated by the internal medullary layer (B21) from the lateral nuclear group (B22).
Subdivision of the latter into a dorsal and a ventral area is less distinct. The entire com- plex is enclosed by a narrow, shell-shaped nucleus, the reticular nucleus of the thalamus (B23), which is separated from the lateral nuclear group by the external medul- lary layer (B24). Ventrally to the thalamus lies the sub- thalamus with the zona incerta (B25) and the subthalamic nucleus (Luys’ body) (B26). The zona incerta is delimited by two myelinated fiber plates, dorsally by Forel’s field H1 (thalamic fasciculus) (B27) and ventrally by Forel’s field H2 (lenticular fasciculus) (B28). Below the subthalamic nucleus appears the rostral pole of the substantia nigra (B29). The floor of the diencephalon is formed by the two mamillary bodies (B30). The mamil- lothalamic fasciculus (Vicq d'Azyr’s bundle) (B31) ascends from the mamillary body to the thalamus.
AB32 Corpus callosum.
A33 Amygdaloid body (amygdala).
B34 Hippocampus.
A35 Anterior commissure.
B36 Medullary stria.
Related Topics