Chapter: Genetics and Molecular Biology: Transposable Genetic Elements
P Element Hopping by Chromosome Rescue
P Element Hopping by Chromosome Rescue
The above-mentioned difficulties with P element
transformation can be overcome by utilizing a property of the cells. Not
surprisingly, parasites have capitalized on the cell’s need for mechanisms to
preserve frag-ments of broken chromosomes. The P elements appear to use this
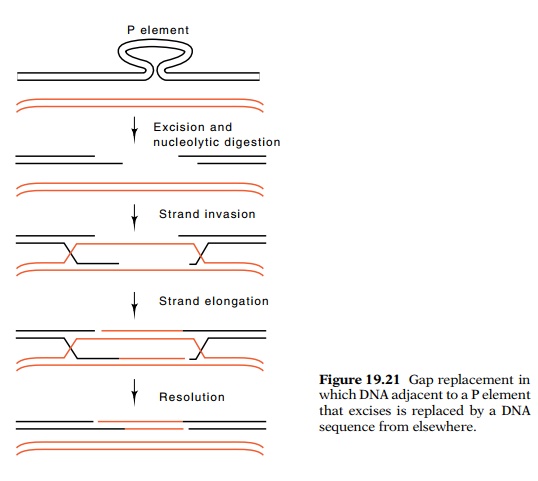
machinery by leaving a gap behind when they excise themselves from one
chromosomal location and insert themselves into another. The cell’s gap repair
or chromosome rescue enzymes then replace the DNA missing from the gap by
making use of a copy of the chromosome that still contains a P element.
Ordinarily, the ends of chromosomes are not
recognized by this repair pathway due to the presence of the telomere
structures. The gaps left by P element excision, however, lack telomeres and
are acted upon by the rescue enzymes.
In the rescue process a small amount of DNA is
digested from the ends by an exonuclease. Then, at each end, one of the two DNA
strands can be further digested so as to leave single-stranded tails. As a
result of RecA-like activity, these single-stranded tails can invade
double-stranded DNA possessing complementary sequences. Subsequent elon-gation
by DNA polymerase and recombination completes the process that removes the gap.
If the P element excision and repair process occurs shortly after chromosome
replication, then the sister chromatids are adjacent to one another and the
centromere has not yet divided. Hence, the most probable template for repair of
a P element excision event is simply the sister chromatid (Fig. 19.21). Thus,
the gap is repaired with another copy of the P element. In toto, a P element has been released with no net change to the
chromosome. Sometimes, however, a P element can excise from the chromosome
before chromosome replica-tion. In this case the template for repair of the gap
can be the homolo - gous chromosome. This other chromosome need not carry a P
element in this position, and therefore the healed chromosome may not contain a
P element.
Consider a P element near or in a region we wish to
modify. If it can be induced to excise, then the repair mechanisms in the cell
can use either the sister chromatid, or the homologous chromosome, or even just
a homologous sequence located on a different chromosome.
Therefore, we can modify the sequence as we desire, and
insert it without a means of expression into the cells. Then we can induce
excision of the P element inactivating our gene. Repair of the resulting gap
can use the specially doctored DNA, and a fraction of the surviving cells will
have replaced a portion of their original gene with our modified sequence.
Hence, a directed mutagenesis step has been performed. This replace-ment does
not require that the P element have carried the entire modified gene, and
further, the DNA sequence we have introduced by this method goes exactly to the
desired chromosome site.
In summary, P elements are useful in genetic
engineering in three basic types of experiments. First, they can be used as
mutagens. They can transpose into genes and inactivate them. Because it is
straightfor-ward to clone the DNA flanking such an insertion event, it is
possible to search for P element transformation events that have inactivated a
desired gene, and then to clone the inactivated gene. A second use of the P
elements is to carry DNA into Drosophila.
Either intact genes can be carried into whatever location a P element hops
into, or fragments of genes can be engineered to replace exactly a region of
DNA already in the chromosome. The third use of P elements is the enhancer trap
experiments that were described. In those experiments, a P element transposes a
reporter gene into the influence region of an enhancer.
Related Topics