Chapter: Genetics and Molecular Biology: Transposable Genetic Elements
An Invertible Segment of Mu Phage
An Invertible Segment of Mu Phage
The split-ended heteroduplexes described in the
preceding section are observed with DNA that is obtained from Mu-infected
cells. Mu DNA obtained instead from an induced lysogen yields heteroduplexes
con-taining not only the split end, but half of them also contain an internal
single-stranded region of 3,000 base pairs that forms a bubble (Fig. 19.11).
This bubble results from the fact that the region, called G, is inverted in about half of the phage.
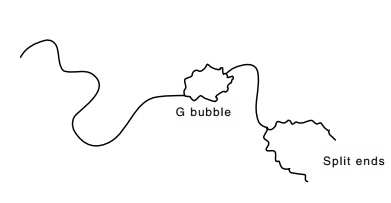
Figure 19.11 Representationof an electron micrograph of a portion of a denatured and rean-nealed Mu phage DNA duplex showing the split ends and G loop
The ability of Mu to invert G can be abolished by a small deletion at the end of G or by a point mutation in the adjacent
region of the DNA called gin. This
point mutation can be complemented, thereby proving that gin encodes a protein that participates in inversion of G. Curiously, phage P1 also contains the
same G loop, and P1 can complement Mu
gin mutant.
The G-loop segment codes for proteins that determine the host range of Mu. Phage induced from lysogens with G in the plus orientation are able to adsorb to Escherichia coli K-12, but not to Citrobacter freundii or Escherichia coli C. When G is in the minus orientation, the abilities of the resultant phage to adsorb to these strains are reversed. This phenomenon provides an explanation of the heteroduplexing experi-ments. Those Mu able to adsorb to Escherichia coli K-12 must be in the plus orientation. Since the rate of inversion is low, the G region of most of the phage in the resulting lysate will still be in the plus orientation, and heteroduplexes prepared from this DNA will not contain G bubbles. Lysogens are different. Although the phage originally entered cells with the G loop in the plus orientation, during the many generations of growth from a single lysogen, G can invert, and eventually about half the cells in the population will contain Mu with G in the plus orientation and half will contain Mu with G in the minus orientation. Induction of the phage in this cell population and preparation of DNA heteroduplexes will yield half the molecules with G bubbles.
The mutants with G stuck in one orientation because of the deletion at the end of G or the point mutation in gin permit a particularly simple
demonstration of the infectivity properties. Mu lysogens with G stuck in the minus orientation yield
phage particles, but these are not infective on Escherichia coli K-12. Mu lysogens containing G stuck in the plus orientation are fully infective when plated on Escherichia coli K-12.
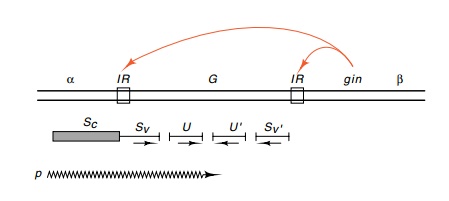
Genetic analysis of the G region of the phage shows that two sets of genes are involved: S and U, and S’ and U’. S
and U are expressed when G is in the plus orientation and S’and U’are expressed when G is
in theminus orientation (Fig. 19.12). The promoter for these genes and the
initial portion of the S gene is
contained in the α region adjacent to G. Thus, G contains a variable portion of S and S’ as well as the intact U
and U’ genes. This is an interesting
example of a situation in which a given DNA sequence can be used to specify different
products.
Related Topics