Chapter: Human Nervous System and Sensory Organs : Telencephalon
Neocortex: Occipital Lobe
Occipital Lobe
The medial surface of the occipital lobe is crossed by the horizontally running cal-carine sulcus (BC1); its deepest point corre-sponds to an eminence on the ventricular wall, the calcar avis (B2). Frontal sections through the occipital lobe clearly show a fiber plate in the white matter, the tapetum(B3). It contains commissural fibers of the corpus callosum that run through the splenium and radiate in an arch into the occipital lobe .
Visual Cortex (Striate Area)
Area 17 (AC4) is the terminal station of the optic radiation and represents thevisual cortex. It lies on the medial surface of the occip-ital lobe and spreads slightly to the convex part at the pole. The cortex lines the calcarine sulcus and extends further to its dor-sal and ventral lips. Area 17 is surrounded by area 18 (A5) (left side of figure, the arrow shows the boundary between areas 17 and 18) and area 19, which represent theopticintegration areas.
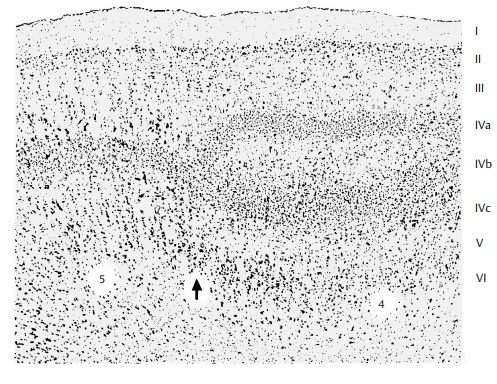
Like all receptive cortical areas, the cortex of area 17 is characterized by reduced pyra-midal layers and well-developed granular layers. The cortex is very thin and set apart from the white matter by a cell-rich layer VI. The internal granule layer (IV) is divided by a zone deficient in cells. In myelin-stained sections, this zone corresponds to Gennari’sline (B6). Because of the striate appearance,the visual cortex is therefore also called striate cortex. In the cell-deficient zone (IVb)are remarkably large cells, the giant stellate cells or Meynert’s cells. The two cellular lay-ers of the internal granular layer (IVa and IVc) contain very small granule cells. These are the layers with the highest cell density within the entire cerebral cortex. Area 18 exhibits a uniform granular layer made up of large granule cells. Area 19 forms a trans-ition to the parietal and temporal cortices.

Functional Organization of the Visual Cortex
Electrophysiological studies on the visual cortex of experimental animals have shown that there are two main types of neurons in the striate area:simple cells and complexcells. Asimple cellreceives impulses from acell group of the retina. Its response is strongest to narrow lines of light, to dark lines against a light background, or to straight lines of light/dark boundaries. Orientation of the lines is crucial: some cells respond only to horizontal lines of light, others only to vertical or oblique lines.
Complex cells also respond to lines of light ofa specific orientation. However, whereas a simple cell becomes excited only by its re-ceptive field, a complex cell responds to mo-bile lines of light that move over the retina: each complex cell is stimulated by a large number of simple cells. It is assumed that the axons of numerous simple cells termi-nate on one complex cell. The internal granular layers consist almost entirely of simple cells, while the complex cells aggre-gate in the external granular layer. More than one half of all neurons in areas 18 and 19 are complex or hypercomplex cells. It is as-sumed that they play a special role in shaperecognition.
Electric stimulation of the visual cortex (area 17) causes the sensation of light sparks or flashes. Stimulation of the parastriate and peristriate areas (areas 18 and 19) is thought to produce figures and shapes; it also causes the gaze to turn (occipital eye field). Eye movements induced by the occipital lobe are purely reflex, in contrast to the volun-tary movements directed by the frontal eyefield.
Functional Organization of the Visual Cortex (continued) (A, B)
Subdivision of the visual cortex into columns.In addition to the structural sub-division into cell layers, the visual cortex shows a functional subdivision into columns; these run vertically to the cell lay-ers and through the entire width of the cor-tex; they are 0.3 – 0.5 mm in diameter. Each column is connected to a defined field of the retina. When the sensory cells of such a field are stimulated, all neurons of the respective column respond. Each column is connected to a peripheral field of only one of the two retinas. In the visual cortex (A1), columns for the right retina alternate with columns for the left retina (ocular dominancecolumns). Hence, the impulses from eachretina are segregated along the entire visual pathway.
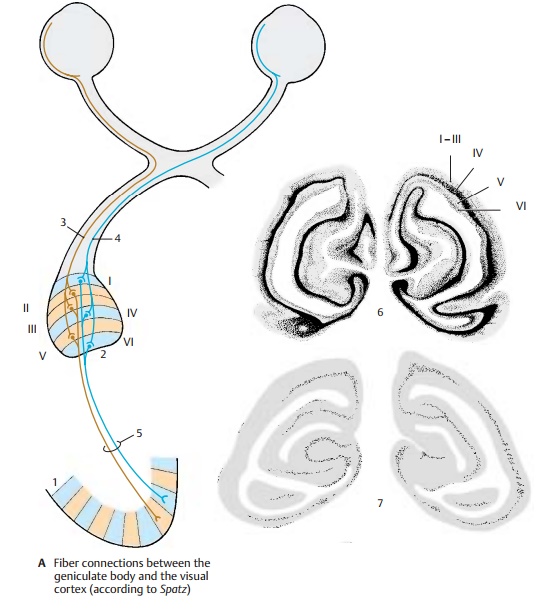
The nerve fibers from the two correspond-ing halves of the retina terminate in the lateral geniculate body: the fibers from the leftretinal halves of both eyes (right halves of the visual fields) terminate in the left geniculate body (A2), and the fibers from the right retinal halves (left halves of the visual fields) terminate in the right genicu-late body. The fibers from corresponding fields of both retinal halves terminate in different cell layers of the geniculate body: the uncrossed fibers (A3) from the ipsi-lateral retina extend to the second, third, and fifth layers, while the crossed fibers (A4) from the contralateral retina run to the first, fourth, and sixth layers. The neurons receiving the optic fibers from correspond-ing points of the two retinas lie along a line that runs through all cell layers (projectioncolumn). Their axons project via the opticradiation (A5) to the visual cortex. Each geniculate fiber is thought to branch exten-sively and to terminate on several thou-sands of stellate cells of cortical layer IV. Fibers conducting the excitation of the ipsi-lateral retina extend to other columns than the fibers for excitation of the contralateral retina.
The organization of the visual cortex into verticalcolumns has been visualized by administer-ing 14C-deoxyglucose to experimental ani-mals and determining the varying distribu-tion of the substance by autoradiography. Excited neurons possess an increased me-tabolism and rapidly take up 14C-deoxyglu-cose, whereas resting cells do not.
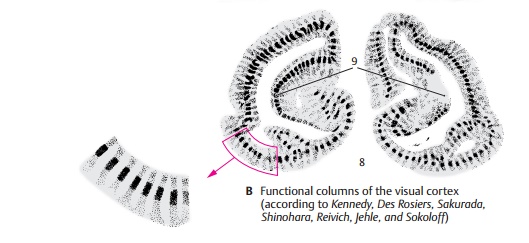
The visual cortex of an experimental animal with both eyes open (rhesus monkey) showed a bandlike distribution of the radioactive label, corresponding to the fa-miliar stratification of cells (B6). The dis-tribution of the label indicated that layers I, II, III, and V have a low glucose content, while layer VI has a higher content and layer IV the highest. When both eyes of the test animal were closed, there were no signifi-cant differences between individual layers; instead, low-grade labeling of relatively uni-form distribution throughout the cortex (B7) was found. If one eye was open while the other one was closed, this method yielded a series of columns running perpen-dicularly to the cell layers and showing in-tensely dark columns alternating with pale ones (B8). The pale columns, where the neurons did not take up the label, repre-sented the retina of the closed eye. The dark columns contained newly incorporated 14C-deoxyglucose because they received input from the retina of the open eye. Again, layer IV was most intensely labeled. The columns were absent in a small area (B9) repre-senting the monocular zones of the retina, namely, the outermost margin of the retina and the blind spot.
In addition to the ocular dominance columns, periodically arranged cell popula-tions have been demonstrated that respond in a characteristic fashion to the orientation of lines in the visual field (orientationcolumns). By demonstrating the enzyme cy-tochromoxidase in sections cut parallel to the cerebral surface, periodically arranged spots have been found that represent neu-rons responding to color stimuli mediated by one eye (color columns).
Related Topics