Chapter: Human Nervous System and Sensory Organs : Telencephalon
Evolution - Telencephalon
Evolution
During primate evolution, the telencephalon has undergone changes similar to those taking place during human embryonic development; it developed late and then overgrew the other parts of the brain. Thus, the cerebellum (A1) is still completely exposed in the brain of primitive mammals (hedgehog), while it becomes more and more covered by the hemispheres of the tel-encephalon during primate evolution.
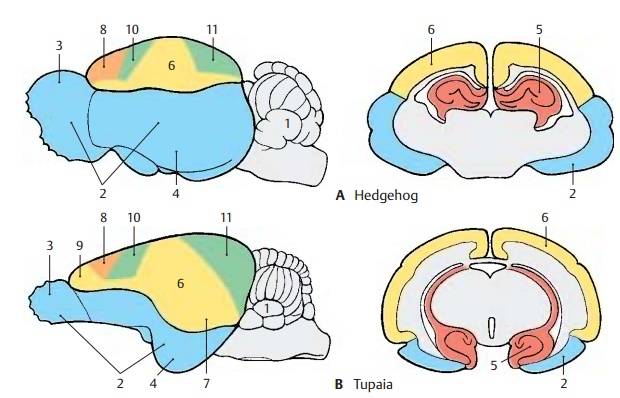
The paleopallium (rhinencephalon) (blue) (A – C2) with olfactory bulb (A – C3) and pir-iform lobe (A–C4) forms the largest part ofthe hemisphere in the primitive mam-malian brain (A), and the archipallium (red) (A – D5) still has its original dorsal position above the diencephalon. These two old components of the hemisphere then be-come overgrown by the neopallium (yellow) (A – D6) during the course of evolution. The paleopallium of prosimians (C) is still of considerable size. In humans (D), however, it becomes displaced deep into the base of the brain and no longer appears in the lateral view of the brain. The archipallium (hippocampus), which lies above the dien-cephalon in the hedgehog (A5), appears as a part of the temporal lobe at the base of the brain in humans (D5). Only a narrow rem-nant remains above the corpus callosum (indusium griseum).
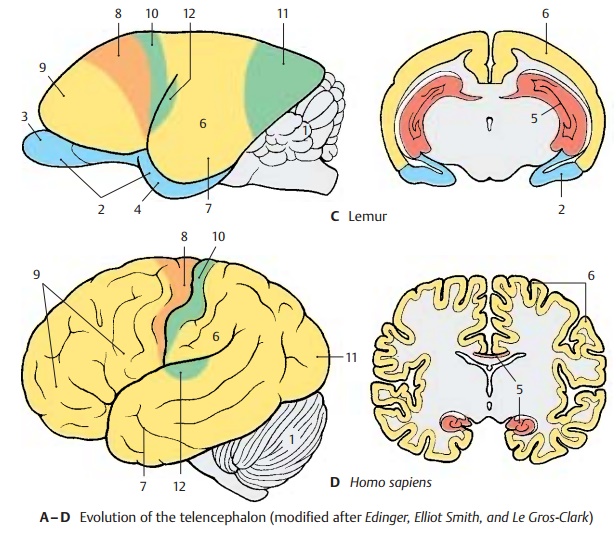
The positional changes largely correspond to the rotation of the hemisphere during embryonic development; they also lead to the formation of the temporal lobe (B – D7). While still absent from the brain of the hedgehog (A), the temporal lobe is already recognized as a ventrally directed projec-tion in the brain of the tree shrew (Tupaia), the most primitive of primates (B). In the prosimian brain (C), a caudally directed temporal lobe has developed that finally turns rostrally in the human brain (D). In ad-dition, sulci and gyri develop in the region of the neopallium. Whereas the neopallium of primitive mammals is smooth (lissen-cephalic brains), a relief of convolutionsdevelops only in higher mammals (gyren-cephalic brains). The development of sulciand gyri considerably enlarges the surface of the cerebral cortex. In humans, only one-third of the cortical surface lies at the sur-face of the hemispheres, two-thirds lie deep in the sulci.
Two types of cortical areas can be distin-guished on the neocortex: the primary areasof origin (light red) and termination areas (green) of long pathways, and between them the secondary association areas (yellow).
The area of origin of motor pathways, the motor cortex (A–D8), constitutes the entirefrontal lobe in the hedgehog. An association area (B – D9) appears for the first time in primitive primates (Tupaia) and achieves extraordinary expansion in the human brain. The termination area of sensory path-ways, the sensory cortex (A – D10), borders caudally on the motor cortex. Owing to the enlargement of the adjacent association area, most of the termination area of the visual pathway, the visual cortex (A – D11), becomes displaced to the medial hemispheric surface in humans. The termi-nation area of the acoustic pathway, the auditory cortex (CD12), becomes displaceddeep into the lateral sulcus (fissure of Syl-vius) by the expansion of the temporal asso-ciation areas. Thus, the association areas ex-pand much more during evolution than the primary areas; they represent the largest part of the neocortex in humans.
Related Topics