Chapter: Human Nervous System and Sensory Organs : Telencephalon
Archicortex
Archicortex
Subdivision and Functional Significance (A – D)
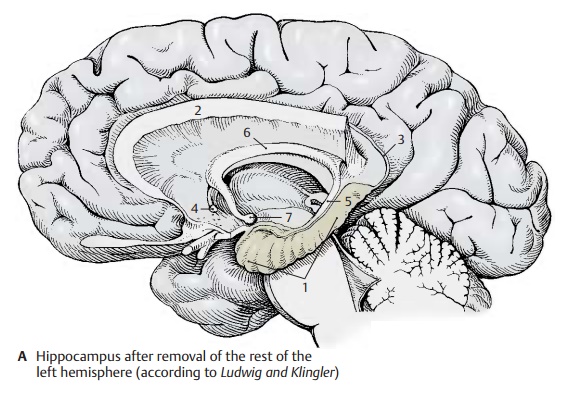
The hippocampus (A – D1) is the main part of the archicortex. It lies at the medial aspect of the temporal lobe in the depth and is largely covered by the parahippocampal gyrus. The left hemisphere has been re-moved in the preparation, showing the cut surface of the corpus callosum (A2) with only the left hippocampus being left intact. The latter looks like a paw with claws, the digitations. The temporal lobe of the righthemisphere in the background illustrates the position of the hippocampus in the tem-poral lobe. The hippocampus extends to the caudal end of the corpus callosum. Here, it becomes reduced to a thin layer of gray matter, the indusium griseum (A3), which ex-tends along the superior surface of the cor-pus callosum to its rostral end in the region of the anterior commissure (A4). Two nar-row fiber bundles, the lateral and the mediallongitudinal striae of Lancisi also run here bilaterally. On the dorsal surface of the hippocampus lies a thick fiber band, the fimbria of hippocampus (A – D5), which separates from the hippocampus beneath the corpus callosum and continues as fornix (A6), arching down to to mamillarybody (A7).
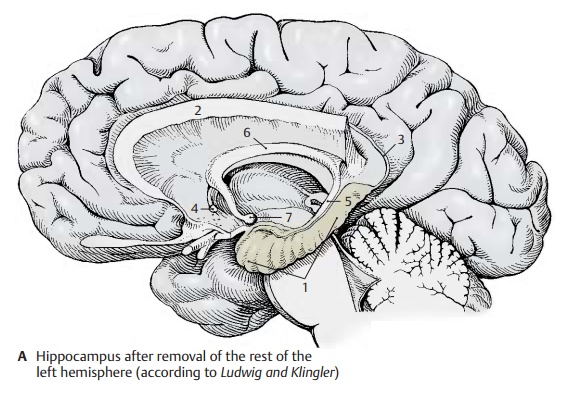
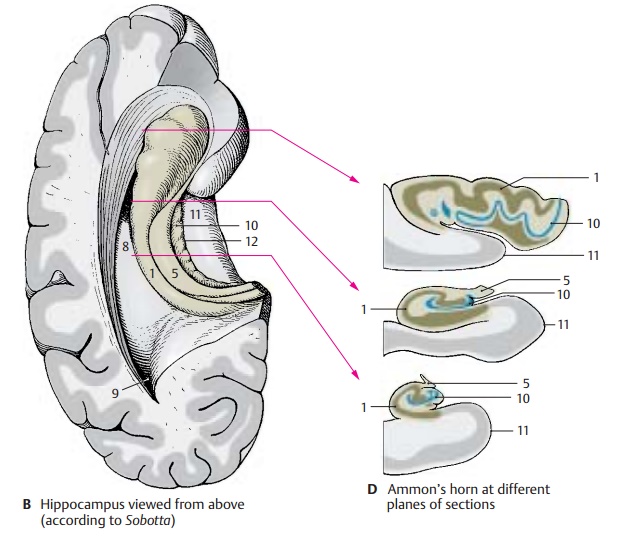
In a horizontal section through the temporal lobe, the inferior horn (BC8) and the poste-rior horn (B9) of the lateral ventricle are ex-posed, and the protrusion of the hippocam-pus into the ventricle is visible. Medially, al-ready at the outer aspect of the temporal lobe, lies the fimbria and, beneath it, the dentate gyrus (fascia dentata) (B – D10), sepa-rated from the parahippocampal gyrus (en-torhinal area) (B – D11) by the hippocampalsulcus (BC12).
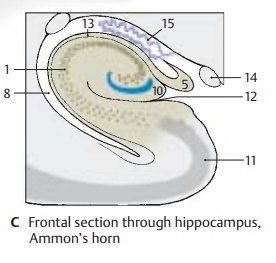
In a frontal section the hippocampal cortex forms a curled band, Ammon’s horn, which protrudes against the ventricle and is covered by a layer of fibers, the alveus (C13). Ammon’s horn shows considerable varia-tions at different planes of section (D)In the past, the hippocampus has been as-signed to the rhinencephalon, but it has no direct relationship with the olfactory sense. In reptiles, which do not have a neocortex, the telencephalon is the highest integration organ. Electrical recordings from the hippo-campus of mammals show that it receives optic, acoustic, tactile, and visceral input, but only a few olfactory impulses. It is an integration organ influencing endocrine, visceral, and emotional processes via its con-nections to the hypothalamus, septal nuclei, and cingulate gyrus. Furthermore, the hip-pocampus plays a major role in processes of learning and memory.
Clinical Note: Bilateral removal of the hippo-campus in humans (treatment of severe epileptic seizures) leads to a loss in memory. While old memories are retained, new information can be remembered only for a few seconds. Such a short-term memory may persist for years. The hippo-campal neurons possess a very low absolute threshold for convulsive discharges. Thus, the hippocampus is of special importance for the origin of epileptic seizures and memory deficits.
C14 Optic tract.
C15 Choroid plexus.
Ammon’s Horn (A)
The hippocampus is subdivided into four parts according to width, cell size, and cell density:
! Field CA1 (A1) contains small pyramidalcells.
! Field CA2 (A2) is characterized by a nar-row, dense band of large pyramidal cells.
! Field CA3 (A3) is characterized by a wideloose band of large pyramidal cells.
! Field CA4 (A4) forms the loosely struc-tured inner zone. Recently, it has been called into question whether a separate CA4 region can be delimited from the CA3 region.
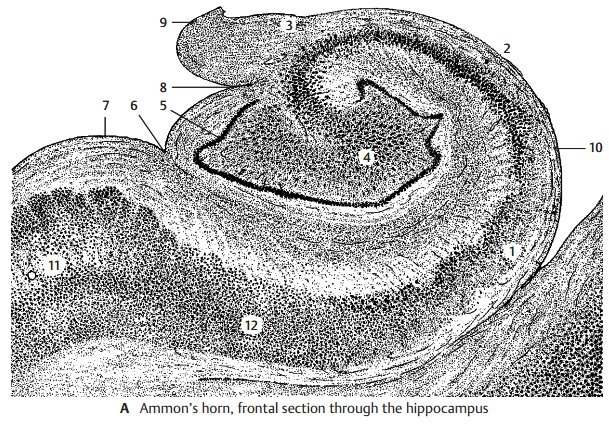
The narrow band of densely packed granule cells of the dentate gyrus (fascia dentata) (A5) surrounds the ending band of pyra-midal cells. The dentate gyrus is fused with the surface of the curled-up Ammon’s horn and appears only partially at the surface of the brain. It is separated by the hippocampalsulcus (A6) from the parahippocampal gyrus (A7) and by the fimbriodentate sulcus (A8) from the fimbria of the hippocampus (A9). The inner layer bordering on the ventricle is the alveus of the hippocampus (A10), in which the efferent fibers collect before leav-ing the hippocampus via the fimbria. The transitional area between Ammon’s horn and the bordering entorhinal cortex (A11) is called the subiculum (A12).
Fiber Connections (B, C)
Afferent Pathways (B)
The fiber bundles from the entorhinal area (B13) are thought to be the most important afferent system, where the pathways from the primary olfactory centers (prepiriform area), from the amygdaloid body, and from various regions of the neocortex terminate. Direct connections between olfactory bulb and hippocampus have not been demon-strated. The fibers from the cingulate gyrus collect in the cingulum (B14) and extend primarily to the subiculum.The fornix (B15) contains bundles from the septal nuclei (B16) but above all fibers fromthe hippocampus and the entorhinal area of the contralateral hemisphere (via the com-missure of the fornix).
Efferent Pathways (B)
Apart from a few fibers leaving the hippo-campus via the longitudinal stria (B17), the fornix contains all other efferent pathways. It is divided into a precommissural part and a postcommissural part. The fibers of the precommissural fornix (B18) terminate in theseptum, in the preoptic area (B19), and in the hypothalamus (B20). The fibers of the postcommissural fornix (B21) terminate in themamillary body (B22) (predominantly in the medial nucleus of the mamillary body), in the anterior thalamic nucleus (B23), and in the hypothalamus. Some fibers of the fornix ex-tend to the central gray matter of the mid-brain.
A large neuronal circuit can be recognized in this system of pathways. Hippocampal im-pulses are conducted via the fornix to the anterior thalamic nucleus. The latter is con-nected with the cingulate gyrus, from where there is feedback via the cingulum to the hippocampus (Papez circuit).
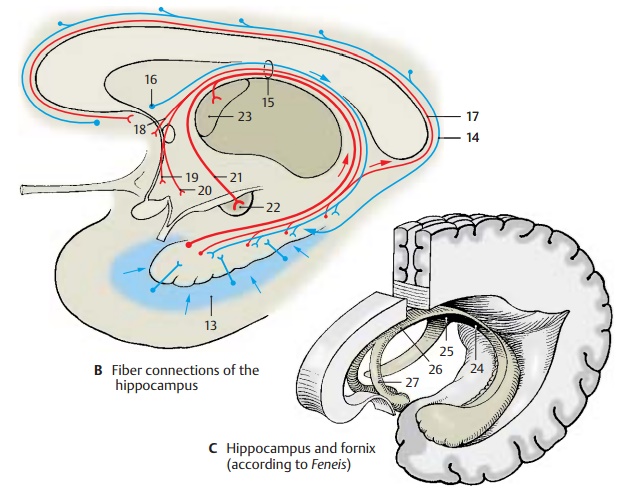
Fornix (C)
At the inferior surface of the corpus callo-sum, the two limbs of the fornix (C24) unite to form the commissure of the fornix (psal-terium) (C25) and the body of the fornix (C26), which then divides again into the two columns of the fornix (C27) above the fora-men of Monro.
Hippocampal Cortex (A, B)
The structure of the archicortex is simpler than that of the neocortex, and its neuronal circuits are therefore easier to elucidate. The hippocampal cortex belongs to those brain regions where inhibitory and excitatory neurons have been identified both histo-logically and electrophysiologically.
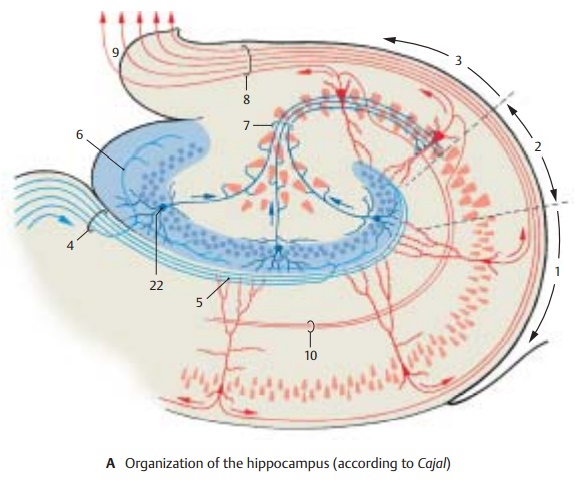
Fields CA1 (A1), CA2 (A2), and CA3 (A3) showdifferences with respect to organization and fiber connections. The majority of afferent fibers enter Ammon’s horn via the perforantpath (A4), and only a few do so via the alveusof hippocampus. They terminate on the dendrites of the pyramidal cells (AB5). Many of the fibers (AB6) extend to the granule cells (AB22) of the dentate gyrus (fascia dentata); their axons, mossy fibers (AB7), too, have synaptic contacts with the dendrites of pyr-amidal cells. However, mossy fibers run only to field CA3; they are absent from fieldsCA1 and CA2.
The pyramidal cells are the efferent el-ements; their axons collect in the alveus (AB8) and leave the cortex through the fim-bria (A9). The axons of the CA3 pyramidalcells give off recurrent collaterals (Schaffercollaterals) (AB10) that form synapses withdendrites of the CA1 pyramidal cells. The efferent fibers running to the septum origi-nate in CA3, the fibers for the mamillary body and the anterior thalamic nucleus originate in CA1. Many of the efferent fibers of the hippocampus, however, run to the subiculum.
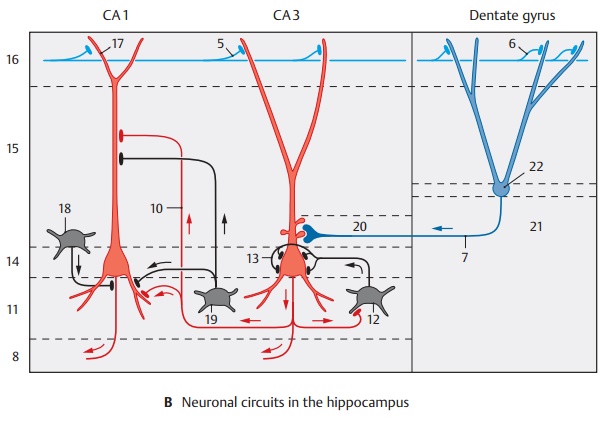
Organization of layers.Ammon’s horn con-sists of the following layers: the alveus (AB8) with the efferent fibers lies inside and is followed by the stratum oriens (B11 ) with the basket cells (B12), the axons of which split up and fill the pyramidal layer with a dense fiber network (B13). The fibers en-velope the pyramidal cell bodies and form synaptic contacts (axosomatic synapses) with them. Basket cells are inhibitory neu-rons that are excited by the axon collaterals of the pyramidal cells and cause pyramidal cell inhibition following pyramidal cell dis-charge. The pyramidal cells form thestratum pyramidale (B14). Their apices areoriented toward the subsequent stratumradiatum (B15), their bases toward the stratum oriens. They send dense dendritictrees in both directions. The long apical den-drite reaches with its branches into the stratum lacunosum-moleculare (B16). In theCA3 region, one can also distinguish a stratum lucidum (B20) where the mossy fibers run.
The afferent fibers originating from different regions run in different layers. Many of the commissural fibers from the contralateral hippocampus extend into the stratum oriens (B11) and the stratum radiatum (B15). The fibers of the entorhinal area (B5) extend into the stratum lacunosum-moleculare (B16) and form contacts with the outermost branches of the apical dendrites (B17). Schaffer collaterals (B10) have contact with distal segments of the apical dendrites of the CA1 pyramidal cells, while the mossy fibers (B7) have contact with proximal seg-ments of the CA3 pyramidal cells. The den-drites of granule cells of the dentate gyrus are contacted in a similar way; entorhinal fibers terminate on distal dendritic seg-ments, while commissural fibers terminate on proximal segments of the dendrites. In addition to the principal cells—pyramidal cells and granule cells—the afferent fibers of the hippocampus also form synaptic con-tacts with inhibitory GABAergic inter-neurons (feed-forward inhibition of princi-pal neurons, p. 35, C). Apart from the basket cells mentioned above (B12), which form axosomatic synapses, GABAergic cells have been found in recent years that form synap-tic contacts on the initial segment of the axon (axo-axonal cells or chandelier cells) (B18) or on the dendrites (B19) of the princi-pal cells. From the course of the fibers and from electrophysiological studies, the fol-lowing impulse flow emerges in the hippo-campus: glutamatergic, entorhinal afferent fibers activate granule cells which, in turn, activate CA3 pyramidal cells via mossy fibers. These then activate CA1 pyramidal cells via Schaffer collaterals (trisynaptic ex-citatory pathway of the hippocampus).
B21 Hilus of dentate gyrus.
Related Topics