Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Fish genetics
Molecular evolution - Fish genetics
Molecular evolution
Access to DNA sequence information has revolutionized the field of phylogenetics. Using models of DNA sequence evolution, researchers can sort out the relationships among fishes from the earliest lineages to the most recent speciation events. Here we review a few of the many breakthroughs in recent years. More examples are accumulating in the scientific literature on a monthly basis. Indeed, it is a good time to be a fish phylogeneticist
The deepest vertebrate radiations: are you a fish?
Fishes arose approximately 530 million years ago, and three deep lineages survive today: the lampreys (Petromyzontiformes), hagfishes (Myxiniformes),and jawed vertebrates (Gnathostomes). Notably, that last category includes cartilaginous fishes (Chondrichthyes), extant bony fishes (Actinopterygii), and tetrapods (amphibians, reptiles, birds, mammals). The fossil record indicates that all these groups arose in the first hundred million years of fish history, however the order in which they arose has been subject to extensive debate.
Takezaki et al. (2003) used over 27 kb of DNA sequence data from 35 nuclear genes to resolve the deepest lineages in the fish tree (Fig. 17.8). Despite fundamental morphological differences, the two jawless fishes (hagfishes and lampreys) appear to be each other’s closest relatives. These data indicate that the cartilaginous fishes diverged next, followed by a bony fishes/tetrapod radiation. Based on molecular studies, the coelacanths appear to diverge near
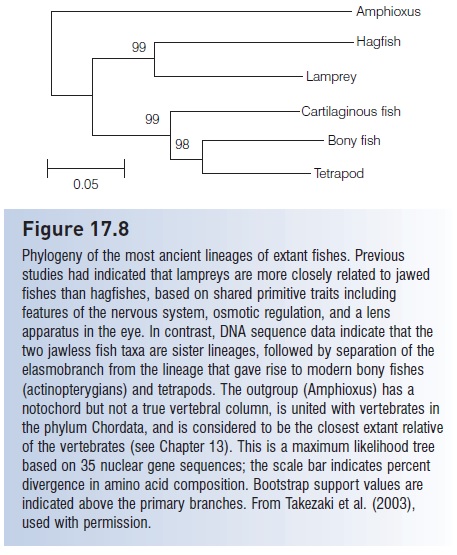
Figure 17.8
Phylogeny of the most ancient lineages of extant fishes. Previous studies had indicated that lampreys are more closely related to jawed fishes than hagfishes, based on shared primitive traits including features of the nervous system, osmotic regulation, and a lens apparatus in the eye. In contrast, DNA sequence data indicate that the two jawless fish taxa are sister lineages, followed by separation of the elasmobranch from the lineage that gave rise to modern bony fishes (actinopterygians) and tetrapods. The outgroup (Amphioxus) has a notochord but not a true vertebral column, is united with vertebrates in the phylum Chordata, and is considered to be the closest extant relative of the vertebrates. This is a maximum likelihood tree based on 35 nuclear gene sequences; the scale bar indicates percent divergence in amino acid composition. Bootstrap support values are indicated above the primary branches. From Takezaki et al. (2003), used with permission.
These studies illustrate two points: First, molecular systematics is especially valuable in cases where the morphology is too divergent (or too similar) to make robust phylogenetic conclusions. Second, the lineage that gave rise to terrestrial vertebrates was the most recent of the major branches in fish history, demonstrating that you, the reader, are really an odd fish.
The history of ray-finned fishes
The most successful modern fishes are the teleosts. However, ray-finned fishes include four additional lineages, known as the ancient actinopterygians. These include polypteriforms (bichirs and reedfish), acipenseriforms (sturgeons and paddlefish), lepisosteids (gars), and Amia calva (Bowfin). How they relate to teleosts, and each other, has been a matter of considerable debate, with systematists proposing almost every possible arrangement of relationships. However, most authorities have identified the polypteriforms as the oldest extant group of ray-finned fishes.
To address the evolutionary history of the ancient and modern ray-finned fishes, Inoue et al. (2003) analyzed entire mtDNA genomes from 12 of the ancient actinopterygians, 14 teleosts, and two elasmobranch outgroups (Fig. 17.9). This extensive DNA sequencing effort, approximately 16.5 kb per species, represents a growing trend in molecular phylogenetics fed by improvements in
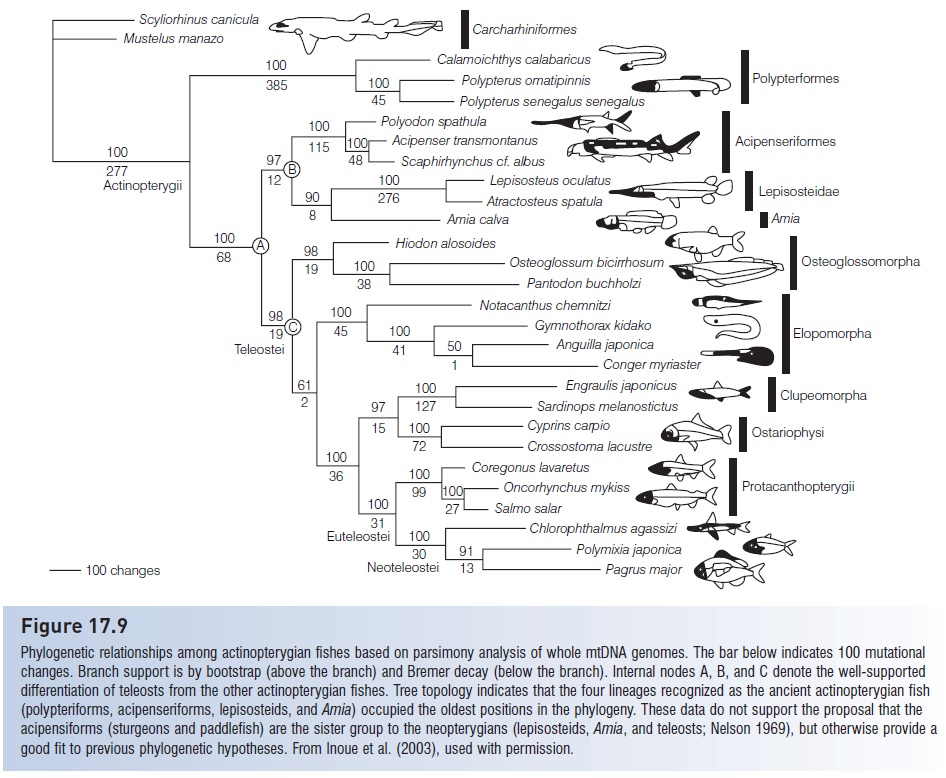
Figure 17.9
Phylogenetic relationships among actinopterygian fishes based on parsimony analysis of whole mtDNA genomes. The bar below indicates 100 mutational changes. Branch support is by bootstrap (above the branch) and Bremer decay (below the branch). Internal nodes A, B, and C denote the well-supported differentiation of teleosts from the other actinopterygian fishes. Tree topology indicates that the four lineages recognized as the ancient actinopterygian fish (polypteriforms, acipenseriforms, lepisosteids, and Amia) occupied the oldest positions in the phylogeny. These data do not support the proposal that the acipensiforms (sturgeons and paddlefish) are the sister group to the neopterygians (lepisosteids, Amia, and teleosts; Nelson 1969), but otherwise provide a good fit to previous phylogenetic hypotheses. From Inoue et al. (2003), used with permission.
In keeping with earlier hypotheses, the polypteriforms appear to be the most ancient of the living ray-finned fishes.
No doubt their persistence into the modern era is aided by unusual adaptations to arid conditions; for example, the bichir live in semipermanent freshwater habitats in Africa and their gas bladder functions as a primitive lung. They can obtain oxygen from air during periods of stagnation and drought, and can move over land to another body of water if their lake or swamp dries up. Based on the mtDNA data, the sturgeons, paddlefish, gars, and Bowfin are a sister lineage to the teleosts.
The phylogeny of higher teleosts (Percomorpha) has been investigated with 100 complete mtDNA sequences, and these data indicate many unexpected relationships, including a phylogenetic affinity between Lophiiformes (goosefish, long assumed to be a primitive teleost) and Tetradontiformes (pufferfishes, long assumed to be among the most advanced teleosts) (Miya et al. 2003). Clearly these findings indicate a rich field for further investigation.
Mapping an evolutionary innovation in parrotfishes
Parrotfishes (Scaridae) are a group of herbivorous fishes that include browsers on seagrass, excavators on hard substrate, and scrapers on coral reefs. Using their beaklike jaws, individuals can consume tons of coral every year, and in undisturbed locations they promote a healthy balance between coral growth and erosion (Bellwood et al. 2003). For this reason they are regarded as ecosystem engineers, essential to ecosystem function.
Several recent studies have attempted to resolve the origin of the unusual “parrot” jaw morphology, in which the teeth are fused to form an efficient tool for removing algae and coral. The oldest known fossil example belongs to one of the seagrass grazers (genus Calotomus), prompting a hypothesis that parrotfish made a gradual transition from the less specialized browsers to the excavators and most recently to the coral scrapers (Bellwood 1994). These evolutionary hypotheses are testable with trait mapping, in
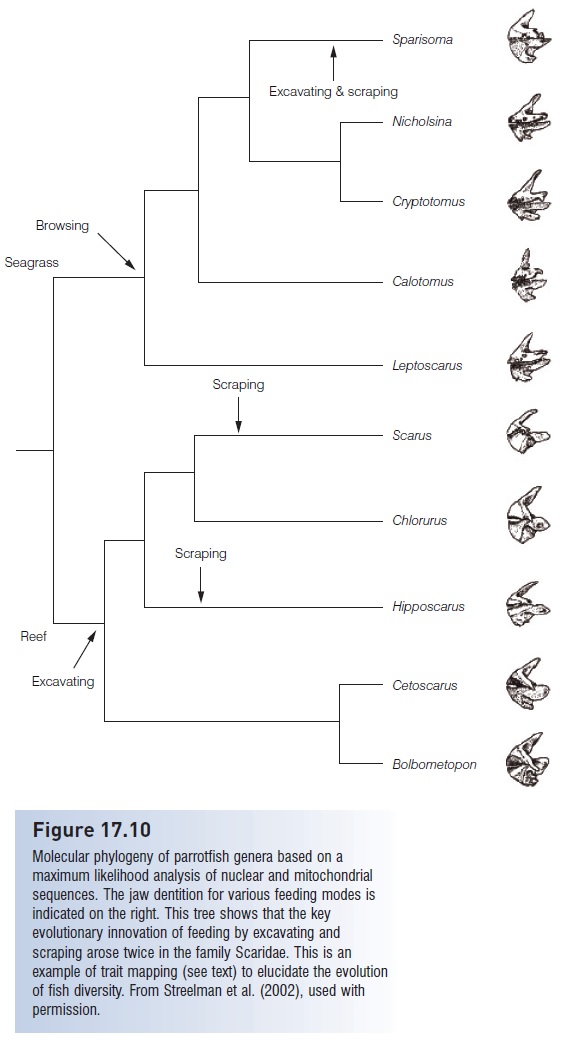
Figure 17.10
Molecular phylogeny of parrotfish genera based on a maximum likelihood analysis of nuclear and mitochondrial sequences. The jaw dentition for various feeding modes is indicated on the right. This tree shows that the key evolutionary innovation of feeding by excavating and scraping arose twice in the family Scaridae. This is an example of trait mapping (see text) to elucidate the evolution
of fish diversity. From Streelman et al. (2002), used with permission.
In this case, browsing, excavating, and scraping can be mapped on a molecular phylogeny of Scaridae, to determine whether the oldest branches in the tree include browsers, and whether the youngest branches include scrapers.
Streelman et al. (2002) addressed this question with 2 kb of DNA sequence, including the nuclear intron Tmo-4C4 and three mtDNA segments (cytochrome b, 12S, and 16S genes) in 16 scarid species and two outgroups (Labridae). The resulting phylogeny (Fig. 17.10) shows an ancient separation between the grazers or browsers versus the excavators and scrapers. The grazer lineage has jaw morphology that is little modified from the ancestral condition, with no fused teeth. The notable exception in the grazer lineage is the genus Sparisoma, which contains browsers, excavators, and scrapers. The Sparisoma species that are excavating or scraping have independently evolved the fused teeth and corresponding jaw morphology. Hence the evolutionary innovation of excavation/scraping arose twice in the family Scaridae. The genus Sparisoma is estimated to be 14–35 million years old based on a molecular clock, providing a timeframe for the independent evolution of the excavating/ scraping adaptation (Bernardi et al. 2000).
Cryptic evolutionary diversity: the case of the bonefishes
Bonefishes (genus Albula) inhabit sand flats in tropical and subtopical habitats, where they are widely sought by anglers because of their high-energy battles at the end of a fishing line. The bonefish was originally described by Linnaeus (1758). Subsequent taxonomic research contributed 23 species names for bonefishes around the world. However as scientific communication improved in the 19th and 20th century, it became apparent that these regional “species” were very similar or indistinguishable. These species were synonymized until most bonefish were recognized as a single species (Whitehead 1986).
The recognition of a single globally distributed bonefish began to unravel when Shaklee and Tamaru (1981) analyzed allozymes in Hawaiian bonefish. They discovered two genetically distinct forms that occupy similar habitats, and that only could be distinguished by careful examination of jaw structure. Subsequent comparisons with mtDNA cytochrome b revealed ancient genetic separations in the genus Albula (d =0.03–0.30), indicating three species in the Caribbean, and three in the East Pacific (Pfeiler et al. 2006; Bowen et al. 2007). Hidaka et al. (2008) discovered subtle morphological differences among Pacific bonefishes, and split one widespread Pacific species into three regional species: A. virgata (a Hawaiian endemic), A. argentea (distributed from the central to West Pacific), and A. oligolepis (West Pacific to Africa). At this writing, there are probably 10 bonefish species, although several have not been formally described (Fig. 17.11). The deepest genetic separation in the genus is between the two sympatric Pacific species A. glossadonta and A. argentea, with an mtDNA cytochrome b sequence divergence of d =0.26–0.30. Based on a molecular clock calibrated for bonefish cytochrome b (1%/Ma), this corresponds to 26–30 million years. It is a remarkable finding that these two fishes, which are identical to the untrained eye and were considered a single species until recently, are five times older than the separation of gorillas and humans.
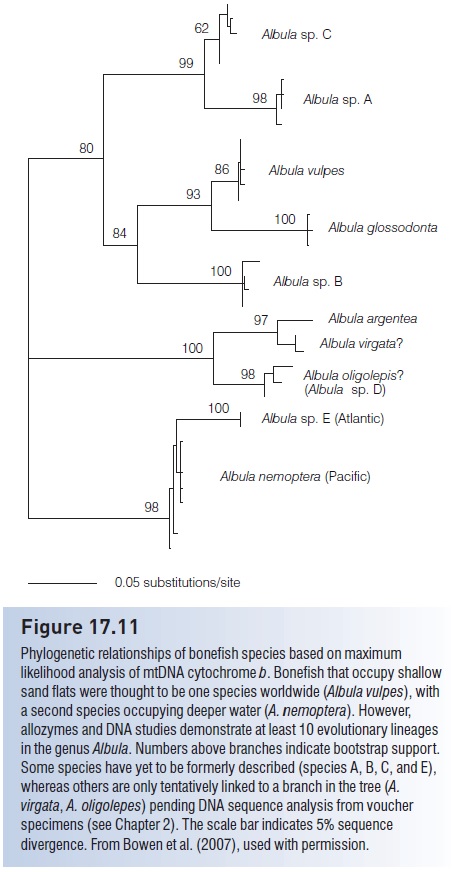
Figure 17.11
Phylogenetic relationships of bonefish species based on maximum likelihood analysis of mtDNA cytochrome b. Bonefish that occupy shallow sand flats were thought to be one species worldwide (Albula vulpes), with a second species occupying deeper water (A. nemoptera). However, allozymes and DNA studies demonstrate at least 10 evolutionary lineages in the genus Albula. Numbers above branches indicate bootstrap support. Some species have yet to be formerly described (species A, B, C, and E), whereas others are only tentatively linked to a branch in the tree (A. virgata, A. oligolepes) pending DNA sequence analysis from voucher specimens. The scale bar indicates 5% sequence divergence. From Bowen et al. (2007), used with permission.
The cichlid radiation of Lake Malawi
In the bonefish example above, genetic studies show that cryptic species can be revealed by mtDNA sequence divergence, most especially in cases where morphological differences are slight or absent. The reverse can also be the case, in which morphological divergence and speciation can outpace mtDNA divergence. The cichlid species flocks of the African Great Lakes have fascinated fish biologists and (more recently) evolutionary geneticists. According to some estimates, Lake Malawi in eastern Africa contains over 600 species, most in the lineage of haplochromine cichlids, with a diversity in form and function that includes eye biters, scale eaters, crab eaters, sediment sifters, plankton eaters, egg robbers, a species that picks parasites off catfish, and one that catches flies near the water’s edge. Taxonomists spent decades sorting these fishes into genera and species, until mtDNA studies upended the whole classification scheme in the 1990s. First, genetic studies demonstrated that the haplochromine cichlids of Lake Malawi are very closely related, d <0.06 in mtDNA sequence comparisons (Albertson et al. 1999). Second, these species descended from a single common ancestor that colonized the lake a few million years ago. Third, many
Fish species flocks exist elsewhere in the world, but none are as diverse as the cichlids of Lake Malawi. What could promote such rapid and extreme diversifi cation? Kocher (2004) describes two factors that seem to promote this process. In the first step, the cichlids move into habitats that require some specialization. Fishes in each habitat will benefit from breeding with similar individuals, to reinforce the genetic and morphological features that allow successful feeding and reproduction. Thisecological selection promotes isolation from cichlids in other habitats, and promotes specialization of feeding morphology and other adaptive traits. The next step is diversifi cation in coloration, a step that can apparently happen on a scale of dozens or hun dreds of generations. Malawi cichlids are nest builders and many are female mouth brooders, behaviors that promote sexual selection wherein females choose a mate based on coloration and behavior. Hence coloration determines which fish interbreed and which ones do not, the foundation of speciation. When the genes for an ecological adaptation are coinherited (perhaps on the same chromosome) with the genes under sexual selection (for distinct coloration), speciation can occur very rapidly. Therefore the composition of the cichlid genome is a third factor that promotes rapid speciation. Kocher (2004) concludes that this plurality of genetic, behavioral, and ecological factors, all of which drive speciation in other organisms, are combined in cichlids to produce the greatest diversity in freshwater fishes. Notably, the cichlid model of speciation does not require geographic isolation (allopatry).
In summary, molecular systematics has revealed much about the history of fishes (and ourselves), and also key points about fish diversity. First, molecular phylogenetics is especially valuable for determining the pattern and pace of evolutionary changes. In the last 20 years the field of systematics has switched from morphology-based trees, to mapping morphological changes on molecular trees. The molecular studies provide a time dimension for these morphological changes, if a calibrated molecular clock is available.
Second, speciation can occur very rapidly, as is the case for African cichlids. Some species of cichlids and other fishes are distinguished by morphology, behavior, and coloration, yet are indistinguishable with mtDNA sequences (Bowen et al. 2006a). A related point is that these rapidly evolving fishes are not isolated by physical barriers, defying the conventional model of allopatric speciation (Wiley 2002; Coyne & Orr 2004). Instead, much of the speciation in fishes seems to occur in adjacent habitats, along ecological rather than geological partitions (Rocha & Bowen2008).
Third, some sister species may be unrecognized because they retain very similar morphology across millions of years, and these hidden species can be revealed with DNA surveys. Cryptic species continue to be discovered, even among the large and well-studied fishes: the numbers of species of ocean sunfishes (genus Mola), goliath groupers (genus Epinephelus), and hammerhead sharks (genus Sphyrna) have all expanded after genetic appraisals (Bass et al. 2005; Quattro et al. 2006; Craig et al. 2008). Very often the genetic difference is accompanied by subtle morphological differences that become apparent upon re-examination. Molecular genetic surveys have also been useful in identifying emerging species, those that seem to be in the process of speciation (McMillan & Palumbi 1995; Campton et al. 2000; Craig et al. 2006).
Finally, the discovery of unrecognized species and cryptic evolutionary diversity can be especially important if these species are scarce, endangered, or heavily exploited. Recall the case of Hawaiian bonefish, a favorite with anglers that was once thought to be a single species, now known to be two species. Consider the implications if the two species, one more common than the other, are managed as a single fishery stock. The less abundant species could be severely depleted without any sign of distress in the overall fishery.
Related Topics