Chapter: Genetics and Molecular Biology: Lambda Phage Integration and Excision
Incorrect Excision and gal and bio Transducing Phage
Incorrect Excision and gal and bio Transducing
Phage
Lambda phage can act as a specialized transducer of
certain host genes. This means that each phage particle in some lambda lines
carries a host gene in addition to essential phage genes. The host gene on the
phage DNA can be brought into an infected cell and either complement a
defective copy of a gene in the host or can recombine with the host copy of the
gene. The process of carrying the DNA across and incorporating it into the
chromosome of the infected cell is called transduction. Because only certain
genes can be transduced by lambda, and because each phage particle in a lysate
of a transducing phage carries a copy of the host gene, the process is called
specialized transduction. Specialized transduction by lambda should be
contrasted to generalized transduc-tion by P1 in which only a small fraction of
the phage particles contain nonphage DNA, only a small fraction of these
contain DNA of any particular bacterial gene, and the DNA in a transducing
particle con-tains no phage DNA.
Transducing
phage once were necessary for the enrichment and ultimate isolation of regions
of DNA involved in regulation of bacterial genes. Also, cells infected with
transducing phage often hypersynthesize the proteins they encode owing to the
presence of multiple phage genomes during much of the phage growth cycle.
Without this gene dosage effect, the proteins could be present in quantities
too low for biochemical study. The enormous value of transducing phage in
re-search on bacterial gene regulation stimulated efforts to develop genetic
engineering techniques so that DNA, proteins, and gene regulation systems from
other organisms could also be studied.
Upon
induction of a lysogen, most prophage excise uneventfully. At a frequency of 10-5
to 10-7, however, a prophage excises incorrectly from the host
chromosome and picks up bacterial DNA bordering the phage integration site.
These incorrect excision events produce transducing phage. One of the joys and
powers of genetics research is that excep-tionally
rare events such as these may be captured and perpetuated for further study.
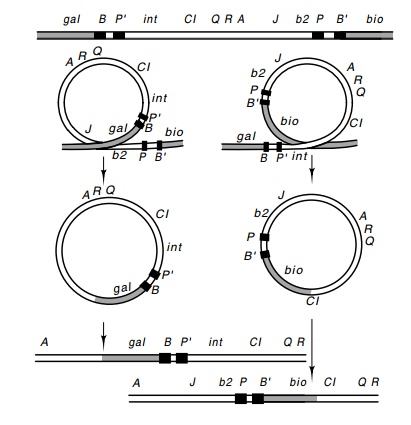
Figure
18.6 Production ofgalandbiotransducing
phage by recombinationevents located to the left or to the right of the end of
an integrated lambda.
When
lambda excises incorrectly from a point other than BOP’ and POB’, bacterial
DNA on one side or the other of the integrated lambdaDNA can be picked up by
the phage (Fig. 18.6). These incorrect excision events could utilize the int and xis products, but more likely use cellular enzymes that break and
rejoin DNA. If the crossover sites are not too far apart for the intervening
DNA to fit within the phage head, the resulting DNA can be encapsidated to form
a viable phage particle. Usually, for packaging to occur some phage genes must
be deleted from the resulting excision product. Looping out to the left
produces a phage with gal genes
substituting for the b2 region. That
is, gal DNA in the phage is flanked
on both sides by lambda genes. Also, attL,
or BOP’, replaces the normal phage
attachment region POP’.
The gal genes are sufficiently far from attλ that phage having picked up gal genes must, of necessity, have left
at least the J gene behind in order
that the transducing phage genome be small enough to be pack - aged in a lambda
coat. These phage are defective because they do not yield viable phage upon
infection of cells. They are propagated by coinfecting them with phage that
provide their missing J gene product.
Such defective gal transducing phage
are called λdgal.
Phage
that excise the other way pick up the biotin genes. These substitute for the
nonessential int, xis, and other early genes under control
of pL. Usually these phage
are not defective and hence are called λpbiofor their ability to form plaques and transduce
biotin genes. Theypossess attR, POB’.
Related Topics