Chapter: Essential Microbiology: Microbial Metabolism
Aerobic respiration
Aerobic
respiration
We shall now examine the fate of the pyruvate produced as the
end-product of glycol-ysis. As we have seen, this depends on whether the
organism in question is aerobic or anaerobic.
You will recall that during glycolysis, NAD+ was reduced to NADH. In order
for glu-cose metabolism to continue, this supply of NAD+ must be replenished; this is
achieved either by respiration or fermentation. Respiration is the term
used to describe those ATP-generating processes, aerobic or anaerobic, by which
oxidation of a substrate occurs, with an inorganic substance acting as the
final electron acceptor. In aerobic respira-tion, that substance is
oxygen; in anaerobic respiration, a
substance such as nitrate or sulphate can fulfil the role.
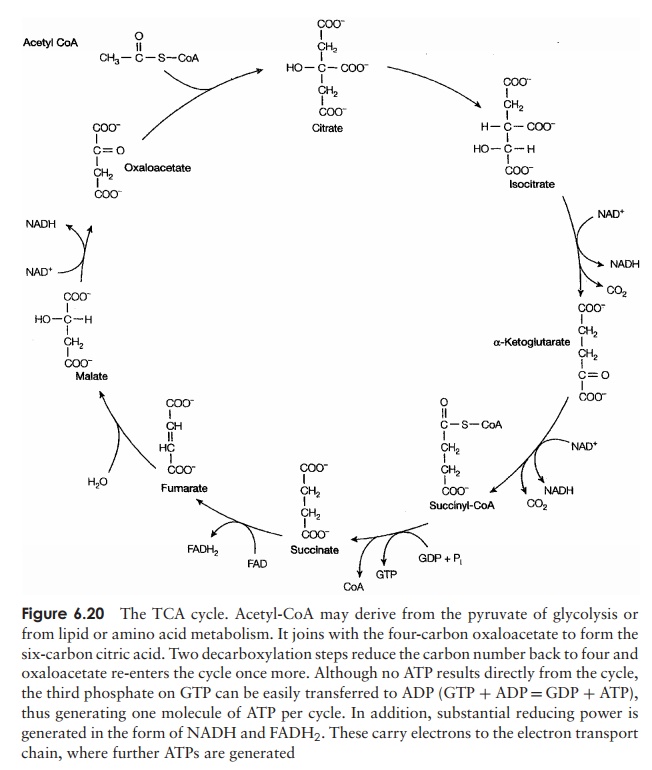
In most aerobic organisms, the pyruvate is completely oxidised
to CO2 and water by entering the tricarboxylicacid
(TCA) cycle, also known as the Krebs
cycle or sim-ply the citric acid
cycle (Figure 6.20). During this cycle, a series of redox reactions result
in the gradual transfer of the energy contained in the pyruvate to coenzymes
(mostly NADH). This energy is finally conserved in the form of ATP by a process
of oxidative phosphorylation. We shall turn our attention to these important
reactionsin due course, but first let us examine the role of the TCA cycle in a
little more detail. Pyruvate does not itself directly participate in the TCA
cycle, but must first be con-verted into the two-carbon compound acetyl-Coenzyme A:
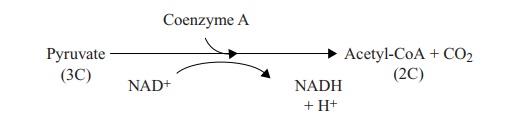
This is an important intermediate, as lipids and amino acids can
also be metabolised into this form, and thereby feed into the TCA cycle. The
main features of the cycle are as follows:
·
each reaction is catalysed by a separate enzyme
·
four of the reactions involve substrate oxidation, with energy,
in the form of electrons, passing to form NADH (mainly) and FADH
·
the two carbons present in acetyl-CoA are removed as CO2
·
one reaction involves the generation of ATP by substrate-level
phosphorylation.
For each ‘turn’ of the citric acid cycle, one molecule of ATP, three molecules of NADH and one molecule of FADH2 are
produced (FADH2 is the reduced form of another coenzyme, FAD). Since
these derive from oxidation of a single acetyl-CoA molecule,
Several of the intermediate molecules in the
TCA cycle also act as precursors in other pathways, such as the synthesis of
amino acids, fatty acids or purines and pyrimidines (see Anabolic metabolism,
below). Other pathways regenerate such intermediates for continued use in the
TCA cycle.
So far, we are a long way short of the 38 molecules of ATP per
molecule of glucose mentioned earlier; we have only managed two ATPs from
glycolysis and a further two
Where do all the rest come from? Most of the
energy originally stored in the glucose molecule is now held in the form of the
reduced coenzymes (NADH and FADH2) produced during glycolysis and
the TCA cycle. This is now converted to no less than 34 molecules of ATP per
glucose molecule by oxidative phosphorylation in the remaining steps in aerobic
respiration (three from each molecule of NADH and two from each of FADH2).

In the final phase of aerobic respiration, electrons are
transferred from NADH and FADH2, via a series of carrier molecules
known collectively as the electrontransport
(or respiratory) chain to oxygen, the terminalelectron
acceptor (Figure 6.21). This in turn is reduced to the molecules of water you
will remember from our overall equation. In procaryotes, this elec-tron
transfer occurs at the plasma membrane, while in eucaryotes it takes place on the
inner membrane of mitochondria. Table 6.2 summarises the locations of the
reactions in the different phases of carbohydrate metabolism.

Related Topics