Chapter: Biotechnology Applying the Genetic Revolution: Molecular Biology of Cancer
Tumor Suppressor Genes or Anti oncogenes
TUMOR-SUPPRESSOR
GENES OR ANTI-ONCOGENES
Tumor-suppressor genes, or
anti-oncogenes, normally suppress cell division. Consequently, it is necessary
to inactivate both copies in order to initiate cancerous growth. A mutation in
just one copy of a tumor-suppressor gene has no effect, that is, these are
recessive mutations. When both copies of a gene have been inactivated by null mutations,
this is known as the nullizygous state. The two most common anti-oncogenes are
Rb and p53 (see later discussion). Almost all human tumors inactivate either Rb
or p53 and often both.
There are two ways to end up with both
copies of a gene inactivated ( Fig. 18.16 ). First, during division of the
cells that form the body, two successive somatic mutations may occur. First one
copy of the gene is inactivated, and then asecond mutation strikes the second
copy of the same gene. Although this seems highly unlikely, it will
occasionally happen given enough time and a sufficiently high number of cells.
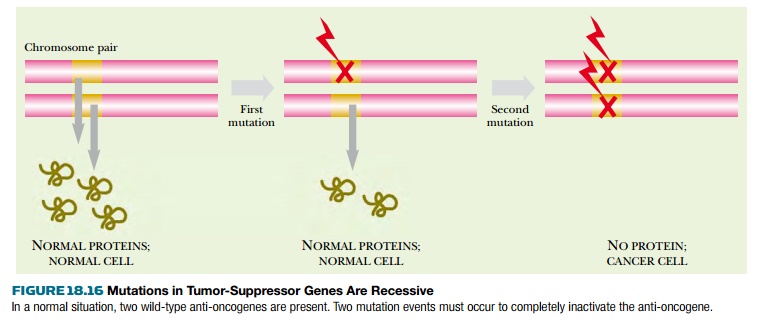
The second route is more frequent. The
first mutation occurs in one copy of the gene in a germline cell of a ancestor.
If this defective copy is passed on, a newborn individual will start life with
one copy of the gene already inactivated in every cell. A somatic mutation that
inactivates the second copy may then occur during cell division. In other
words, one defective allele is inherited and the other is acquired by somatic
mutation. Individuals who inherit a single defective anti-oncogene do not
always develop cancer. What they inherit is an increased chance of doing so.
This scheme applies to a dozen or more
tumorsuppressor genes. Some of them are involved in a wide range of cancers,
such as the retinoblastoma (Rb) gene , originally discovered and named for a
rare cancer of the retina of the eye, but also involved in many other tumors.
Other anti-oncogenes are very tissue specific and inactivation of both copies
triggers cancer of a particular organ, such as the Wilms’ tumor gene
responsible for a kidney cancer.
Anti-oncogenes encode proteins whose normal role is to inhibit growth or prevent cell division. Some signals that promote growth and division come from outside the cell. In other words, hormones circulating in the blood control growth and development. Some proteins encoded by anti-oncogenes are involved in detecting these external hormonal signals and blocking their effect on cell division ( Fig. 18.17 ). An example is the PTEN protein that antagonizes insulin signaling and thus restricts cell growth and protein synthesis.
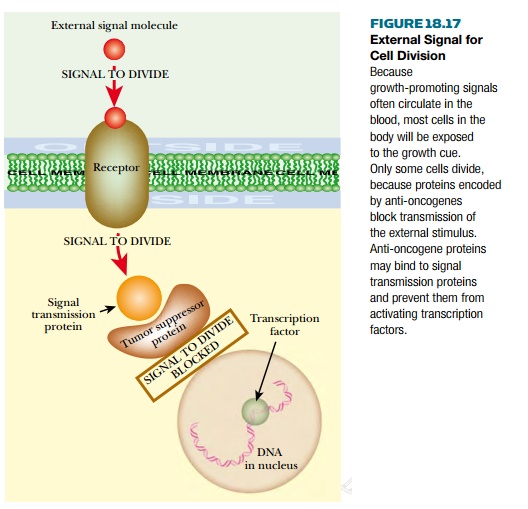
More recently, evidence has accumulated that in addition to external control, many somatic cell lines are also preprogrammed. They have a preset number of allowed cell divisions. An internal “generation clock” of some sort counts off the number of permitted divisions left, and when it reaches zero, growth and division stop. Some anti-oncogenes are part of this system, and when they are defective the cell fails to stop dividing. Indeed, many of the proteins encoded by anti-oncogenes are DNA binding proteins, often with zinc fingers.
Related Topics