Chapter: Human Neuroanatomy(Fundamental and Clinical): The Diencephalon
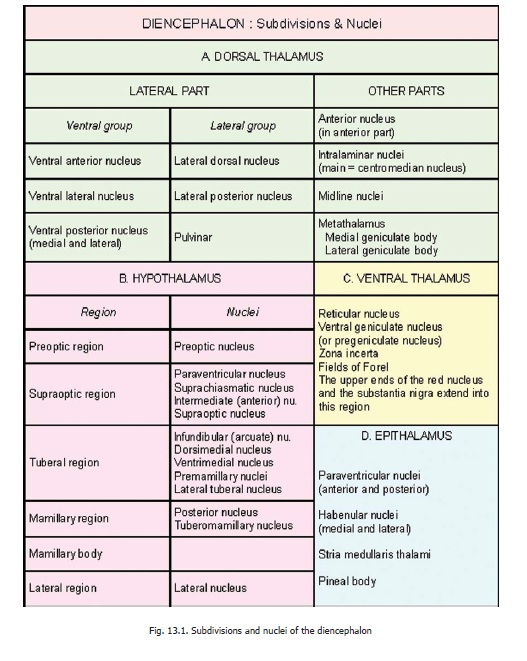
The Thalamus (Dorsal Thalamus) - The Diencephalon
The Thalamus (Dorsal Thalamus)
The thalamus (or dorsal thalamus) is a large mass of grey matter that lies immediately lateral to the third ventricle. It has two ends (or poles), anterior and posterior; and four surfaces, superior, inferior, medial and lateral.
The anterior end (or pole) lies just behind the interventricular foramen (Fig. 8.6). The posteriorend (or pole) is called the pulvinar. It lies just above and lateral to the superior colliculus. Thepulvinar is separated from the geniculate bodies by the superior brachium quadrigeminum.
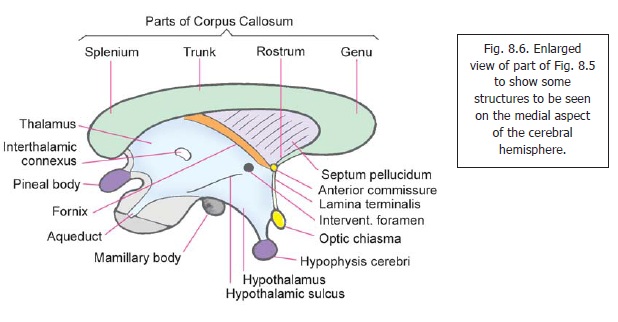
The medial surface forms the greater part of the lateral wall of the third ventricle, and is lined by ependyma. The medial surfaces of the two thalami are usually interconnected by a mass of grey matter called the interthalamic adhesion (connexus). Inferiorly, the medial surface is separated from the hypothalamus by thehypothalamic sulcus. This sulcus runs from the interventricular foramen to the aqueduct. The lateral surface of the thalamus is related to the internal capsule which separates it from the lentiform nucleus (Fig. 13.2). The superior (or dorsal) surface of the thalamus is related laterally to the caudate nucleus from which it is separated by a bundle of fibres called the stria terminalis, and by the thalamostriate vein. The thalamus and the caudate nucleus togetherform the floor of the central part of the lateral ventricle (Fig. 13.2). The medial part of the superior surface of the thalamus is, however, separated from the ventricle by the fornix, and by a fold of piamater called the tela choroidea.
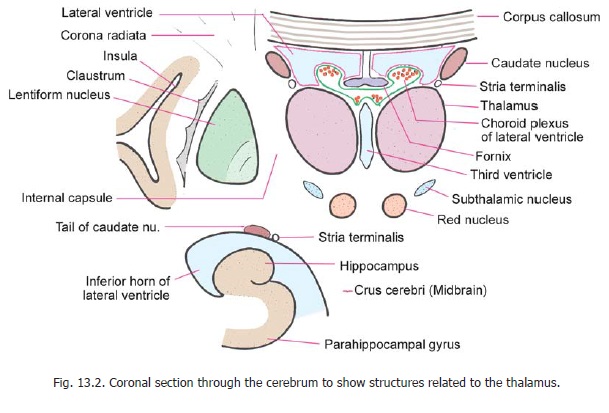
At the junction of the medial and superior surfaces of the thalamus the ependyma of the third ventricle is reflected from the lateral wall to the roof. The line of reflection is marked by a line called the taenia thalami. Underlying it there is a narrow bundle of fibres called the stria medullaristhalami (not to be confused with the stria medullares present in the floor of the fourth ventricle).The inferior surface of the thalamus is related to the hypothalamus anteriorly (Fig. 8.5), and to the ventral thalamus posteriorly (Fig. 13.2). The ventral thalamus separates the thalamus from the tegmentum of the midbrain.
Internal Structure of the Thalamus
The thalamus consists mainly of grey matter. Its superior surface is covered by a thin layer of white matter called the stratum zonale; and its lateral surface by a similar layer called the externalmedullary lamina.
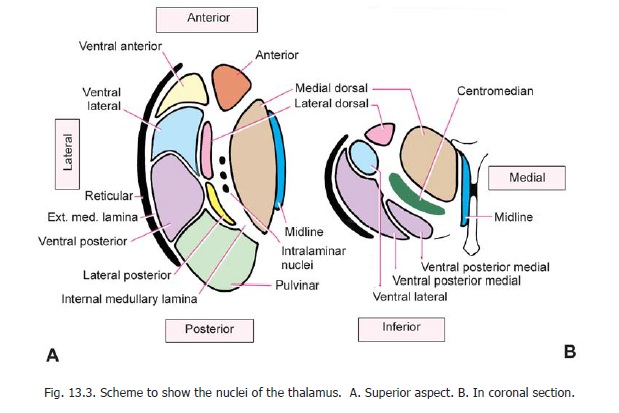
The grey matter of the thalamus is subdivided into three main parts by a Y-shaped sheet of white matter which is called the internal medullary lamina (Fig. 13.3). This lamina is placed vertically. It divides the thalamus into a lateral part, a medial part, and an anterior part situated between the two limbs of the ‘Y’.
A number of nuclei can be distinguished within each of these parts. Only the more important of these are listed below
(a) Nuclei in the anterior part
a. A number of nuclei can be distinguished, but we shall refer to them collectively as the anteriornucleus.
(b) Nuclei in the medial part
a. The largest of these is the medial dorsal nucleus. It is divisible into a magnocellular part (anteromedial) and a parvocellular part (posterolateral).
(c) Nuclei in the lateral part
The nuclei in the lateral part can be subdivided into a ventral group and a lateral group.
The nuclei in the ventral group are as follows (in anteroposterior order).
1. Ventral anterior nucleus.
2. Ventral lateral nucleus (also called the ventral intermediate nucleus).
3. Ventral posterior nucleus, which is further subdivided into a lateral part called theventralposterolateral nucleus, and a medial part called the ventral posteromedial nucleus (Fig. 13.3B).
The nuclei of the lateral group are as follows (in anteroposterior order).
1. Lateral dorsal nucleus (or dorsolateral nucleus).
2. Lateral posterior nucleus.
3. Pulvinar.
(d)Other thalamic nuclei
In addition to the above the thalamus contains the following nuclei.
1. The intralaminar nuclei are embedded within the internal medullary lamina. There are several nuclei in this group. The most important of these is the centromedian nucleus (Fig. 13.3B).
2. The midline nuclei consist of scattered cells that lie between the medial part of the thalamus and the ependyma of the third ventricle. Several nuclei are recognised.
3. The medial and lateral geniculate bodies (traditionally described under metathalamus) are now included as part of the thalamus.
The reticular nucleus, earlier described as part of the dorsal thalamus, is now regarded as part of the ventral thalamus.
Connections of the Thalamus
Afferent impulses from a large number of subcortical centres converge on the thalamus (Fig. 13.4). Exteroceptive and proprioceptive impulses ascend to it through the medial lemniscus, the spinothalamic tracts, and the trigeminothalamic tract. Visual and auditory impulses reach the lateral and medial geniculate bodies respectively. Sensations of taste are conveyed to the thalamus through solitariothalamic fibres. Although the thalamus does not receive direct olfactory impulses they probably reach it through the amygdaloid complex. Visceral information is conveyed from the hypothalamus,
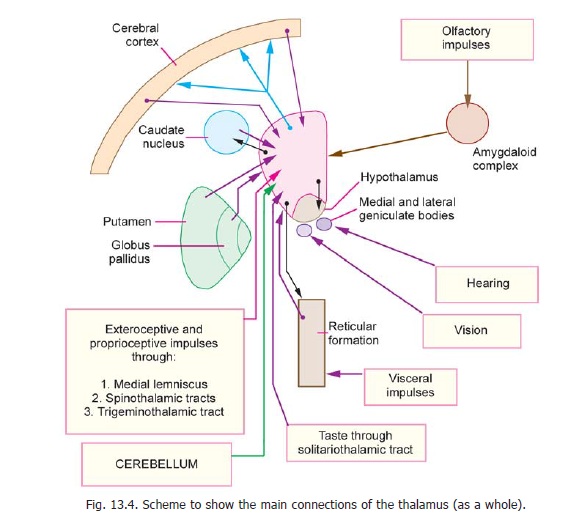
and probably through the reticular formation. In addition to these afferents, the thalamus receives profuse connections from all parts of the cerebral cortex, the cerebellum, and the corpus striatum. The thalamus is, therefore, regarded as a great integrating centre where information from all these sources is brought together. This information is projected to almost the whole of the cerebral cortex through profuse thalamocortical projections. The areas of cerebral cortex receiving fibres from individual thalamic nuclei are shown in Fig. 13.5a. Thalamocortical fibres form large bundles that are described as thalamic radiations or as thalamic peduncles. These radiations are anterior(or frontal), superior (or dorsal), posterior (or caudal), and ventral. Efferent projections from the thalamusalso reach the corpus striatum, the hypothalamus, and the reticular formation. Besides its integrating function, the thalamus is believed to have some degree of ability to perceive exteroceptive sensations, specially pain.
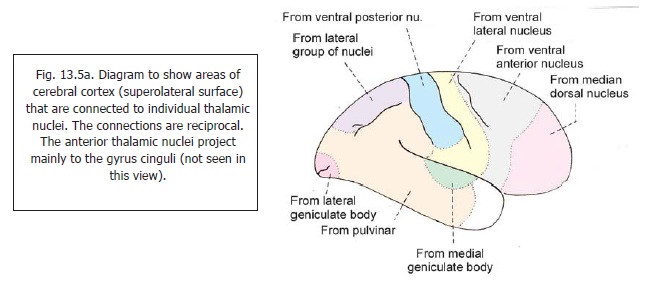
Keeping these facts in mind we may now consider the connections of individual thalamic nuclei. These connections are numerous, and many are controversial. The description that follows is confined to better known connections.
Connections of Ventral Group of nuclei
1. From a clinical point of view the most important connections of the thalamus are those of theventral posterior nucleus. We have seen that this nucleus is divisible into ventral posterolateraland ventral posteromedial parts (that are sometimes mentioned as separate nuclei). This nucleus
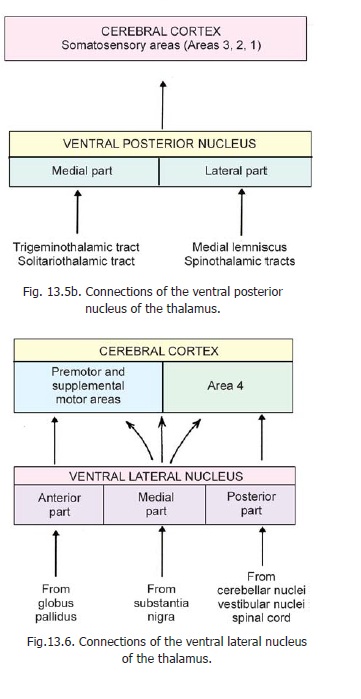
receives the terminations of the major sensory pathways ascending from the spinal cord and brainstem (Fig. 13.5b). These include the medial lemniscus, the spinothalamic tracts, the trigeminal lemniscus and the solitariothalamic fibres carrying sensations of taste. Within the nucleus fibres from different parts of the body terminate in a definite sequence. The fibres from the lowest parts of the body end in the most lateral part of the nucleus. The medial lemniscus and spinothalamic tracts carrying sensations from the limbs and trunk end in the ventral posterolateral part; while the trigeminal fibres (from the head) end in the ventral posteromedial part, which also receives the fibres for taste. Different layers of cells within the nucleus respond to different modalities of sensation.
All the sensations reaching the nucleus are carried primarily to the sensory area of the cerebral cortex (SI, areas 3,2,1) by fibres passing through the posterior limb of the internal capsule (superior thalamic radiation). They also reach the second somatosensory area (SII) located in the parietal operculum (of the insula).
2. Theventral lateral nucleusis divisible intoanterior, posterior and medial parts. Each part has distinct connections (Fig. 13.6).
The anterior part of the nucleus receives fibres from the globus pallidus and projects to the premotor and supplemental motor areas of the cerebral cortex.
The medial part receives afferents from the substantia nigra; and gives off a diffuse projection reaching many areas of the cerebral cortex.
3. The connections of theventral anterior nucleusare largely unknown. It probably receivesfibres from the globus pallidus and cerebellar nuclei, and sends efferents to the cerebral cortex (parietal lobe, anterior part). Its functions are unknown.
connections of lateral group of nuclei
1. The lateral dorsal nucleus receives impulses from the superior colliculus (Fig. 13.7). Efferent projections reach the cingulate gyrus, the parahippocampal gyrus, and parts of the hippocampal formation. Some fibres reach the cortex of the parietal lobe.
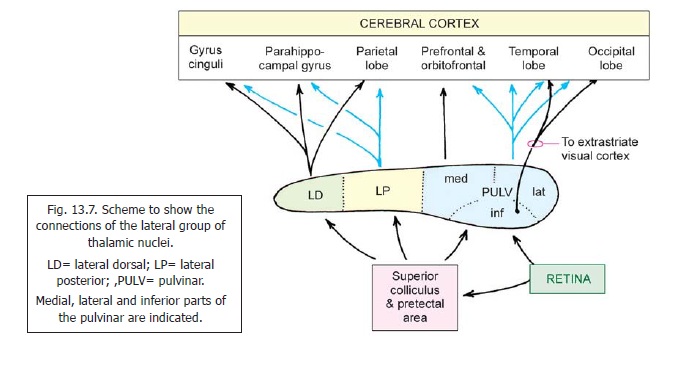
2. The lateral posterior nucleus receives fibres from the superior colliculus. Efferents reach the cerebral cortex of the superior parietal lobule. They also reach the cingulate and parahippocampal gyri.
3. Thepulvinaris divisible into medial, lateral, and inferior parts. (An anterior, or oral, part is also described). All parts receive fibres from the superior colliculus. The inferior part of the pulvinar also receives direct fibres from the retina. It is believed that the entire retina is represented in this part. Efferents from the pulvinar project to extrastriate visual areas in the occipital and parietal lobes; and to visual association areas in the posterior part of the temporal lobe. The inferior part of the pulvinar is thus a centre in an extrageniculate pathway from the retina to the cerebral cortex. However, the projections of the pulvinar to non-visual areas of the cerebral cortex suggest complex functions that may involve recognition and memory.
Connections of other thalamic nuclei
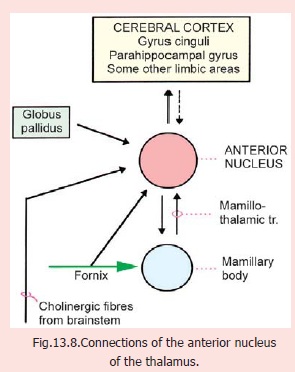
1. Theanterior nucleusreceives fibres fromthe mamillary body through the mamillothalamic tract. Efferent fibres project to the gyrus cinguli (areas 23, 24, 32).Integrity of this pathway is necessary for recent memory. Some other connections of the nucleus are shown in Fig. 13.8. Anterior thalamic nuclei are probably involved in functions involving attention and memory.
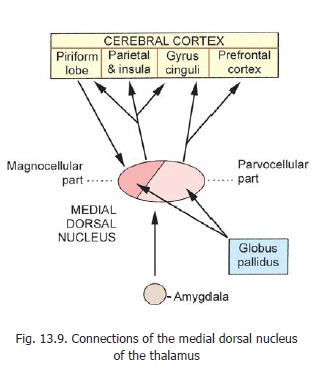
2. Themedial dorsal nucleusis divisibleinto a smaller magnocellular part (placed anteromedially), and a larger parvocellular part (placed posterolaterally). The connections of the nucleus are shown in Fig. 13.9. The nucleus is involved in controlling emotional states. Damage to the nucleus leads to decrease in anxiety, tension and aggression. These functions are similar to those of the prefrontal cortex.
3. Intralaminar nuclei: There are severalnuclei in this group divided into subgroups, anterior and posterior. The posterior subgroup includes the large centromedian nucleus. The nuclei of this group receive inputs from the body through collaterals of spinothalamic tracts (Fig. 13.10). Fibres are also received from the reticular formation, the cerebellar nuclei, and the substantia nigra. The centromedian nucleus receives many fibres from the globus pallidus.
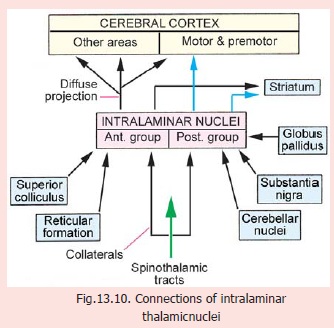
Efferents from intralaminar nuclei reach the cerebral cortex. Those from the anterior subgroup are diffuse reaching many parts of the cortex. Those from the posterior group project to the motor, premotor and supplemental motor areas. Efferents also reach the striatum. Functions of these nuclei are not known.
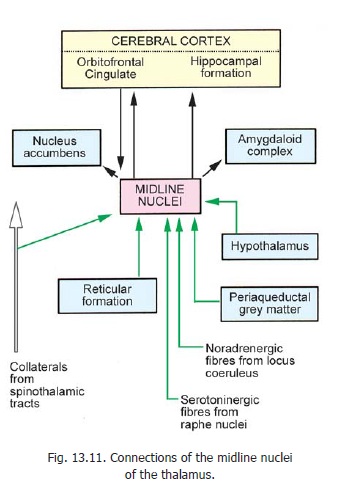
4. Themidline nucleiconsist of several small groups of neurons (but there is controversyregarding the groups to be included under this heading). The connections of the nuclei (shown in Fig. 13.11) are mainly with the limbic system. Afferents include noradrenergic, serotoninergic and cholinergic bundles ascending from the brainstem. The midline nuclei probably play a role in memory and arousal.
The intralaminar, midline and reticular nuclei, have in the past, been grouped together as nonspecificthalamic nuclei. In the past they have been regarded as part of the ascending reticular activatingsystem which is believed to be responsible for maintaining a state of alertness. They have been described as receiving afferents from the reticular formation (mainly gigantocellular nucleus and ventral reticular nucleus of the medulla; caudal reticular nucleus of pons) and projecting to all parts of the cerebral cortex. Recent investigations have shown, however, that there is no validity in dividing thalamic nuclei into specific and unspecific.
Medial & Lateral Geniculate Bodies
The medial and lateral geniculate bodies are small oval collections of grey matter situated below the posterior part of the thalamus, lateral to the colliculi of the midbrain (Fig. 13.12). Each mass of grey matter is bent on itself, hence the term ‘geniculate’. Traditionally, the geniculate bodies have been grouped together under the heading meta-thalamus; but because of functional relationships they are now included in the dorsal thalamus.
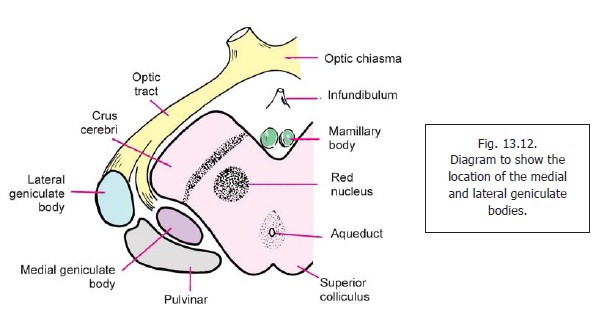
The Medial Geniculate Body
The medial geniculate body is a relay station on the auditory pathway. Medial, ventral and dorsal nuclei are described within it.
The medial geniculate body receives fibres of the lateral lemniscus either directly, or after relay in the inferior colliculus (Fig. 13.13). These fibres pass through the brachium of the inferior colliculus. Fibres arising in the medial geniculate body constitute the acoustic radiation. The acoustic radiation passes through the sublentiform part of the internal capsule to reach the acoustic areas of the cerebral cortex. Some other connections of the medial geniculate body are shown in Fig. 13.13.
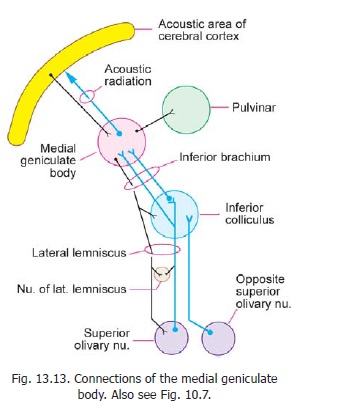
Each medial geniculate body receives impulses from the cochleae of both sides. It also receives fibres from the auditory area of the cerebral cortex. These fibres form part of the descending acousticpathway.
Different neurons in the ventral nucleus of the medial geniculate body respond to different frequencies of sound (tonotopic organisation). The ventral nucleus projects to the primary auditory cortex. The neurons in the dorsal nucleus do not show tonotopic organisation. They project to auditory areas around the primary auditory area.
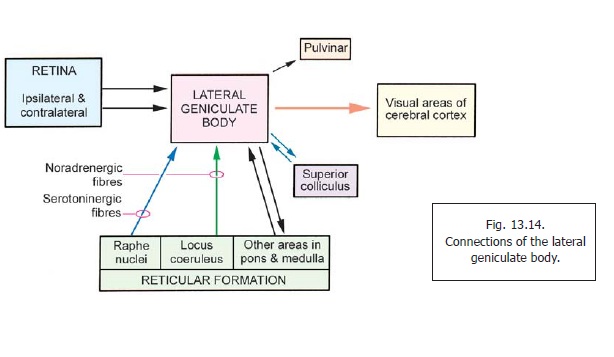
The Lateral Geniculate Body
The lateral geniculate body is a relay station on the visual pathway. It receives fibres from the retinae of both eyes (Fig. 13.14). Efferents arising in this body constitute the optic radiation which passes through the retrolentiform part of the internal capsule to reach the visual areas of the cerebral cortex. Details of the visual pathway (including the representation of retinal quadrants in the lateral geniculate body) are described.
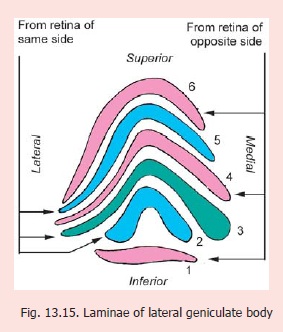
Sections through the lateral geniculate body show that its grey matter is partially split to form six lamellae separated by nerve fibres (Fig. 13.15). These layers are numbered one to six from ventral to dorsal side. Laminae one, four and six receive fibres from the retina of the opposite side; while laminae two, three and five receive fibres from the retina of the same side. The fibres from different parts of the retinae project to specific parts of the lateral geniculate body, the whole of the retina being represented (visiotopic organisation). In turn these project to specific areas of the primary visual cortex (area 17).
Apart from retinal fibres the lateral geniculate body receives fibres from the primary visual cortex, and from extrastriate visual areas. It also receives fibres from the superior colliculus, and from the reticular formation of the pons and medulla.
Noradrenergic fibres reach it from the locus coeruleus, and serotoninergic fibres from raphe nuclei (midbrain).
The various fibres reaching the lateral geniculate body form complicated synapses. The synaptic areas often form glomeruli somewhat similar to those in the cerebellum. In addition to neurons that relay retinal impulses to the visual cortex, the lateral remain confined within it (Golgi type II neurons) and participate in the synapses.
The lateral geniculate body has traditionally been regarded as a simple relay station on the visual pathway. However, both anatomical and physiological evidence shows that considerable interaction of various impulses occurs in the geniculate body. However, the predominant view at present is that impulses from the right and left retinae are not integrated in the lateral geniculate body. Such integration takes place only in the cerebral cortex.
Related Topics