Chapter: Human Neuroanatomy(Fundamental and Clinical): The Diencephalon
The Hypothalamus - The Diencephalon
The Hypothalamus
The hypothalamus is a part of the diencephalon. As its name implies it lies below the thalamus. On the medial side, it forms the wall of the third ventricle below the level of the hypothalamic sulcus. Laterally, it is in contact with the internal capsule, and (in the posterior part) with the ventral thalamus (subthalamus). Posteriorly, the hypothalamus merges with the ventral thalamus, and through it with the tegmentum of the midbrain. Anteriorly, it extends up to the lamina terminalis, and merges with certain olfactory structures in the region of the anterior perforated substance. Inferiorly, the hypothalamus is related to structures in the floor of the third ventricle. These are the tuber cinereum, the infundibulum, and the mamillary bodies, which are considered as parts of the hypothalamus.
Subdivisions of the Hypothalamus
For convenience of description the hypothalamus may be subdivided, roughly, into a number of regions (Figs. 13.16, 13.17). Some authorities divide it (from medial to lateral side) into three zones which are as follows.
a) Periventricular zone.
b) Intermediate zone.
c) Lateral zone.
The periventricular and intermediate zones are often described collectively as the medial zone and we will follow this practice here. The column of the fornix lies between the medial and lateral zones. (The mamillothalamic tract and the fasciculus retroflexus also lie in this plane).
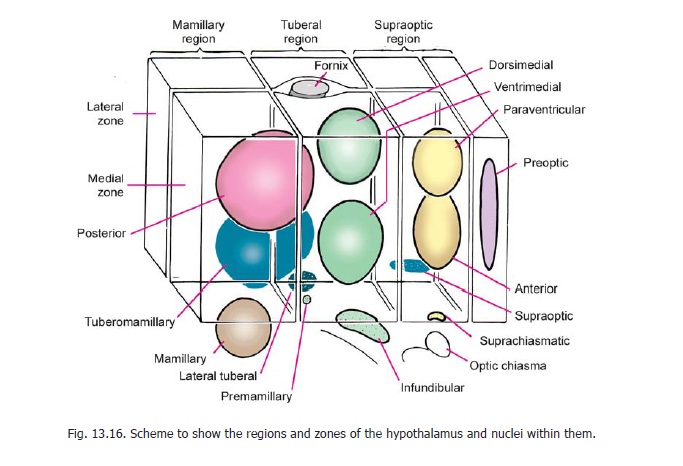
The hypothalamus is also subdivided anteroposteriorly into four regions. These are as follows.
a) Thepreoptic regionadjoins the lamina terminalis.
b) The supraoptic (or chiasmatic) region lies above the optic chiasma.
c) The tuberal (or infundibulotuberal) region includes the infundibulum, the tuber cinereum and the region above it.
d) The mamillary (or posterior) region consists of the mamillary body and the region above it.
The preoptic region differs from the rest of the hypothalamus in being a derivative of the telencephalon. (The lamina terminalis also belongs to the telencephalon).
Hypothalamic nuclei
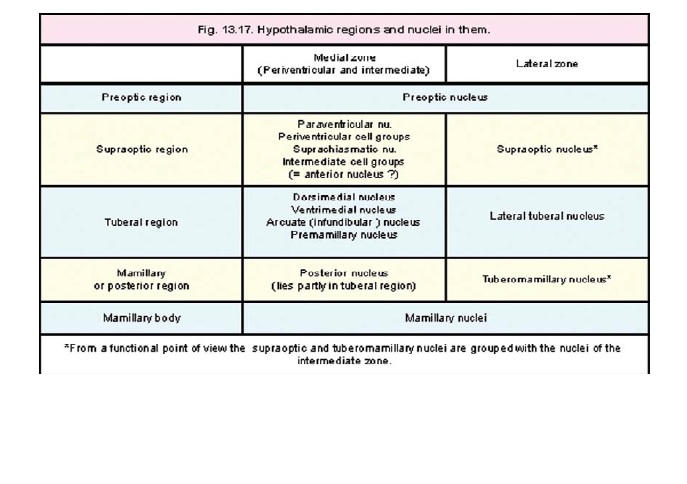
The entire hypothalamus contains scattered neurons within which some aggregations can be recognized. These aggregations, termed the hypothalamic nuclei, are as follows (Figs. 13.16 to 13.18).
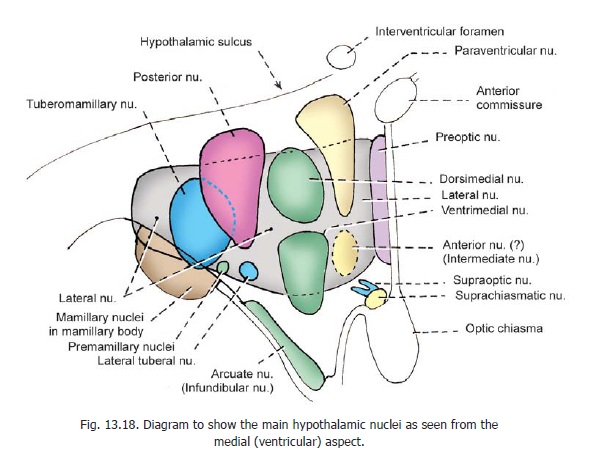
1. The preoptic nucleus extends through the periventricular, intermediate, and lateral zones of the preoptic part.
2. The mamillary nuclei lie within the mamillary body.
The remaining nuclei of the hypothalamus lie either in the periventricular, intermediate, or lateral zones. As the boundaries of nuclei are ill defined, different authors have allocated nuclei differently to these zones. To lessen confusion on this account we will consider the periventricular and intermediate zones collectively as the medial zone.
It has also been found that groupings of hypothalamic nuclei based on functional considerations do not necessary follow groupings according to the regions they lie in. The classification that follows is based on traditional teaching, and some comments on functional groupings are made below.
Nuclei in the medial zone
3. The paraventricular nucleus, and
4. thesuprachiasmatic nucleus, lie in the supraoptic region.
5. The arcuate (or infundibular) nucleus lies in the tuberal region.
6. The posterior nucleus extends into both the tuberal and mamillary regions.
7. The anterior nucleus occupies the supraoptic region.
8. The dorsimedial nucleus, and
9. the ventrimedial nucleus lie in the tuberal part, which also contains small aggregations of cells that constitute
10. thepremamillary nuclei.
Nuclei in the lateral zone
11. The lateral zone contains a diffuse collection of cells that extend through the supraoptic, tuberal and mamillary regions. These cells constitute the lateral nucleus.The lateral zone also contains the following nuclei.
12. The supraoptic nucleus lies in the supraoptic region (just above the optic tract).
13. The tuberomamillary nucleus extends into the tuberal and mamillary regions.
14. Small aggregations of neurons in the tuberal region constitute the lateral tuberal nuclei
Some additional comments on grouping of hypothalamic nuclei
1. On the basis of functional considerations the supraoptic and tuberomamillary nuclei (of the lateral zone) are grouped along with nuclei in the medial zone.
2. The anterior nucleus is not mentioned in some recent texts. Instead a group of intermediatenuclei is described in this situation.
Neuromediators in the Hypothalamus
As many as 25 neuromediators have been located in the hypothalamus, and their distribution has been mapped out. The details are beyond the scope of this book.
Connections of the Hypothalamus
The hypothalamus is concerned with visceral function and is, therefore, connected to other areas having a similar function. These include the various parts of the limbic system, the reticular formation, and autonomic centres in the brainstem and spinal cord (Figs. 13.19, 13.20). Apart from its neural connections, the hypothalamus also acts by releasing secretions into the blood stream, and into CSF.
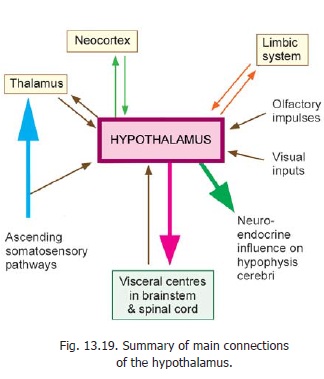
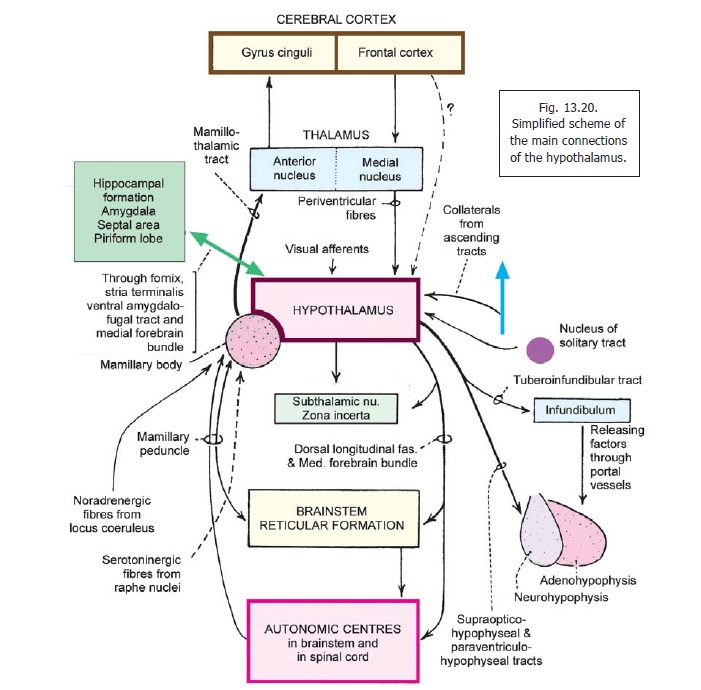
Afferent Connections
1. The hypothalamus receives visceralafferents (including those of taste) through the spinal cord and brainstem. The exact pathways are not known. They probably pass through the reticular formation and consist of several relays.
Many of these fibres pass through a bundle called the mamillary peduncle. Other fibres pass through a bundle called the dorsal longitudinal fasciculus. Fibres from the tegmentum of the midbrain also reach the hypothalamus through the medial forebrain bundle.
2. Afferents from the nucleus of the solitary tract carry taste impulses (and other visceral sensations).
3. Somatic afferents reach the hypothalamus through collaterals of major ascending tracts.
4. The hypothalamus receives afferents from several centres connected to olfactory pathways, and to the limbic system.
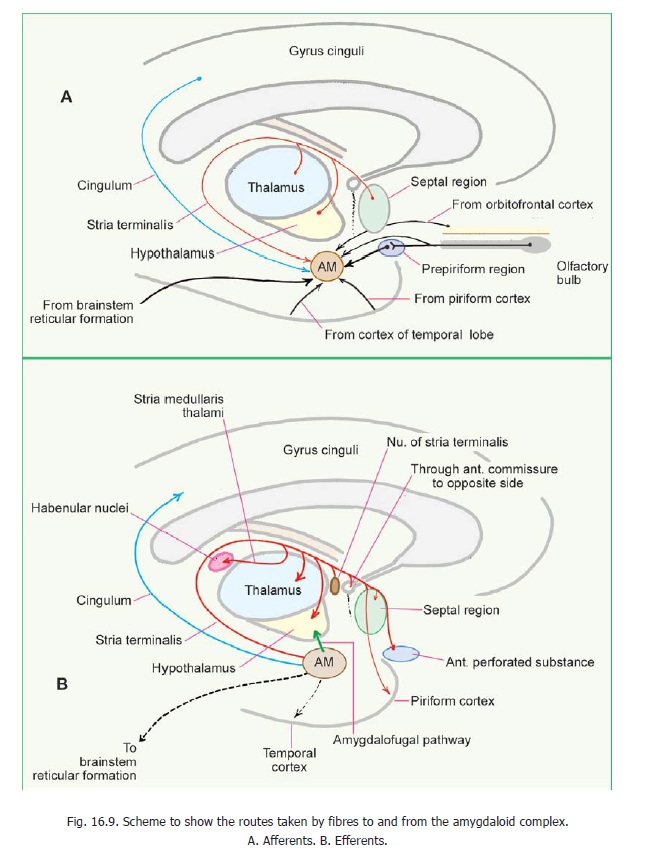
These are the anterior perforated substance, the septal nuclei, the amygdaloid complex, the hippocampus and the piriform cortex. Many of these fibres reach the hypothalamus through the medial forebrain bundle. Fibres from the hippocampus travel through the fornix. Some fibres from the amygdaloid complex pass through the stria terminalis, and some through the ventral amygdofugal tract (Fig. 16.9).
Olfactory impulses are received after relay in the nucleus accumbens (in the ventral thalamus, near the anterior perforated substance); and through septal nuclei.
Visceral impulses reach the suprachiasmatic nucleus.
The locus coeruleus is connected to the hypothalamus through diffuse noradrenergic fibres. The raphe nuclei project to the hypothalamus through serotoninergic fibres. Some cholinergic and dopaminergic fibres also reach the hypothalamus.
5. Cortico-hypothalamic fibres: In addition to fibres from the piriform cortex (mentionedabove) the hypothalamus is believed to receive fibres from the cortex of the frontal lobe. Some of these are direct. Others relay in the thalamus (medial dorsal and midline nuclei) and reach the hypothalamus through periventricular fibres (so called because they travel just subjacent to the ependyma). The gyrus cinguli may influence the hypothalamus indirectly through the hippocampal formation. Some fibres from the orbital cortex may reach the hypothalamus through the medial forebrain bundle.
6. The hypothalamus also receives fibres from the subthalamic nucleus, and the zona incerta.
Efferent Connections
1. The hypothalamus sends fibres to autonomic centres in the brainstem and spinal cord. Centres in the brainstem receiving such fibres include the nucleus of the solitary tract, the dorsal nucleus of the vagus, the nucleus ambiguus, and the parabrachial nucleus. Fibres descending to the spinal cord end in neurons in the intermediolateral grey column. It also sends fibres to the hippocampal formation, the septal nuclei, the amygdaloid complex, and the tegmentum of the midbrain, and autonomic centres in the brainstem and spinal cord. These fibres pass through the same bundles that convey afferent fibres from these centres.
2. Fibres from the mamillary body pass through the mamillothalamic tract to reach the anterior nucleus of the thalamus. New fibres arising here project to the gyrus cinguli. Fibres from the mamillary nuclei also reach the subthalamic region and the tegmentum. (through the mamillo-tegmental tract).
3. Fibres from the hypothalamus project widelyto the neocortex. They play a role in maintaining cortical arousal.
Control of hypophysis cerebri by the hypothalamus.
Neurons in some hypothalamic nuclei produce bioactive peptides that are discharged in the neighbourhood of capillaries; or in some cases into CSF. The process of the production of such bioactive substances by neurons (as distinct from release of neurotransmitters at synapses or efferent nerve endings) is referred to as neurosecretion.
The peptides (hormones) are synthesised by the neuron and are seen as membrane bound granules. Each peptide is associated with a neurophysin which binds the peptide. The granules move along axons to their terminations. Neurosecretory material in cell somata and in axons can be demonstrated by suitable staining. Release of peptide is triggered by activity of the neuron (and is associated with influx of Ca++ ions at the terminals). The released peptides are absorbed into blood. The release of peptides depends on inputs to the neurons concerned. Neurosecretory cells are influenced by temperature and osmolarity of blood, and by concentrations in it of hormones and of nutrients.
Control of Neurohypophysis
It is now known that vasopressin (antidiuretic hormone) and oxytocin, associated with the neurohypophysis, are really neurosecretory products synthesised in the paraventricular and supraoptic nuclei of the hypothalamus. The cells concerned are large (magnocellular secretion). Axons of the paraventricular nucleus descend towards the supraoptic nucleus as the paraventri-culohypophyseal tract (Fig. 13.21). Theyjoin axons arising from the supraoptic nucleus to form the supraoptico-hypophysealtract. The axons of the tract pass down intothe infundibulum and from there into the neurohypophysis. Here the axons branch
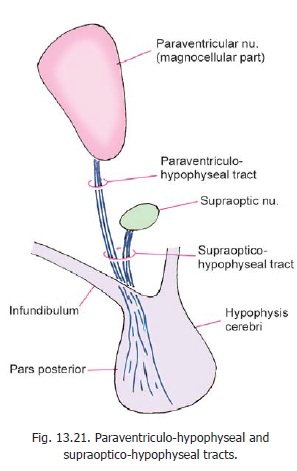
profusely and end in relation to capillaries around which they release their secretion.
The neurons producing vasopressin and oxytocin are distinct. Although both types of neurons are found in each nucleus, the supraoptic nucleus is believed to mainly produce vasopressin, and the paraventricular nucleus is believed to mainly produce oxytocin.
Control of the adenohypophysis by the hypothalamus
The hypothalamus controls secretion of hormones by the adenohypophysis by producing a number of releasing factors. Axons of cells in the infundibular (arcuate) nucleus end in the median eminence and infundibulum. They travel through the tuberohypophyseal tract which also receives fibres from several other hypothalamic nuclei. The axon terminals of the fibres in these tracts are closely related to capillaries in the region. The cells of the arcuate (infundibular) nucleus produce releasing factors that travel along their axons and are released into the capillaries. These capillaries carry these factors into the pars anterior of the hypophysis cerebri through the hypothalamo-hypophysealportal system. In the pars anterior these factors are responsible for release of appropriate hormones.It may be noted, however, that in the case of some hormones their secretion is inhibited by such factors.
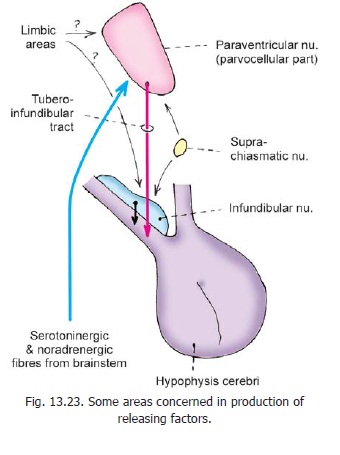
In addition to the arcuate nucleus some other hypothalamic nuclei are also involved in production of releasing factors. The nuclei and the releasing factors produced by them are shown in Fig. 13.22. Fig. 13.23 shows some pathways through which the production of releasing factors may be controlled.

Fibre bundles associated with the hypothalamus
In considering the connections of the hypothalamus reference has been made to a number of fibre bundles not described earlier. These are briefly considered below.
Medial forebrain bundle
The medial forebrain bundle begins in the anterior olfactory areas. It runs caudally through the lateral zone of the hypothalamus, to reach the tegmentum of the midbrain. It contains both ascending and descending fibres. Cholinergic, noradrenergic and dopaminergic fasciculi pass through this bundle. Various other neuromediators have also been demonstrated in relation to this bundle.
Mamillary peduncle
The mamillary peduncle is a bundle of fibres that connects the tegmentum of the midbrain to the mamillary body. The fibres in it carry visceral impulses to the hypothalamus (Fig. 13.20).
Dorsal longitudinal fasciculus
The dorsal longitudinal fasciculus (of Schultz) runs vertically within the brainstem, in close relation to the central grey matter. It is made of unmyelinated fibres and is, therefore, also called the unmyelinated longitudinal fasciculus. It connects centres in the midbrain, pons, and the upperpart of the medulla with the hypothalamus and thalamus. The fibres in it are both ascending and descending.
Central tegmental tract
This tract descends through the midbrain. It lies lateral to the medial longitudinal fasciculus, dorsolateral to the red nucleus. When traced upwards it ascends through the ventral thalamic (subthalamic) region to reach the lentiform nucleus. Some fibres even reach the motor cortex. Traced downwards the tract extends to the medulla where it reaches the inferior olivary complex, and the reticular formation.
In the brainstem, most the fibres in the tract are those arising in the parvocellular part of the red nucleus. These fibres descend to the inferior olivary complex. The central tegmental tract connects the motor cortex and red nucleus to the reticular formation and the inferior olivary complex.
Some fibres in the tract are serotoninergic, noradrenergic or cholinergic. The noradrenergic fibres arise from the nucleus coeruleus.
Functions of the Hypothalamus
The hypothalamus plays an important role in the control of many functions that are vital for the survival of an animal. In exercising such control the hypothalamus acts in close coordination with higher centres including the limbic system and the prefrontal cortex, and with autonomic centres in the brainstem and spinal cord. The main functions attributed to the hypothalamus are as follows.
Regulation of eating and drinking behavior
The hypothalamus is responsible for feelings of hunger and of satiety, and this determines whether the animal will accept or refuse food. It has been observed that stimulation of the lateral zone of the hypothalamus stimulates hunger while stimulation of the medial zone produces satiety. The lateral zone is also responsible for thirst and drinking. Based on such studies a feeding centre has been described in the lateral hypothalamic nucleus, and a satiety centre in the ventromedial nucleus.
Correlate this with the fact that some neurons in the hypothalamus are sensitive to osmolarity (preoptic nucleus), glucose content and fatty acid content of blood.
Regulation of sexual activity and reproduction
The hypothalamus controls sexual activity, both in the male and female. It also exerts an effect on gametogenesis, on ovarian and uterine cycles, and on the development of secondary sexual characters. These effects are produced by influencing the secretion of gonadotropic hormones by the hypophysis cerebri.
Control of autonomic activity
The hypothalamus exerts an important influence on the activity of the autonomic nervous system, and thus has considerable effect on cardiovascular, respiratory and alimentary functions. Sympathetic activity is said to be controlled, predominantly, by caudal parts of the hypothalamus; and parasympathetic activity by cranial parts, but there is considerable overlap between the regions concerned.
Emotional behavior
The hypothalamus has an important influence on emotions like fear, anger and pleasure. Stimulation of some areas of the hypothalamus produces sensations of pleasure, while stimulation of other regions produces pain or other unpleasant effects.
Control of endocrine activity
The influence of the hypothalamus in the production of hormones by the pars anterior of the hypophysis cerebri, and the elaboration of oxytocin and the antidiuretic hormone by the hypothalamus itself, have been described above. Through control of the adenohypophysis, the hypothalamus indirectly influences the thyroid gland, the adrenal cortex, and the gonads.
Response to stress.
Through control over the autonomic nervous system, and hormones, the hypothalamus plays a complex role in the way a person responds to stress.
Temperature regulation
Some neurons in the preoptic nucleus of the hypothalamus act as a thermostat to control body temperature. When body temperature rises or falls, appropriate mechanisms are brought into play to bring the temperature back to normal.
Biological clock
Several functions of the body show a cyclic variation in activity, over the twenty four hours of a day. The most conspicuous of these is the cycle of sleep and waking. Such cycles (called circadianrhythms) are believed to be controlled by the hypothalamus, which is said to function as abiological clock. The suprachiasmatic nucleus is believed to play an important role in this regard. Lesions of the hypothalamus disturb the sleep-waking cycle.
Related Topics