Chapter: Essentials of Anatomy and Physiology: Digestive System
Stomach
STOMACH
Anatomy of the Stomach
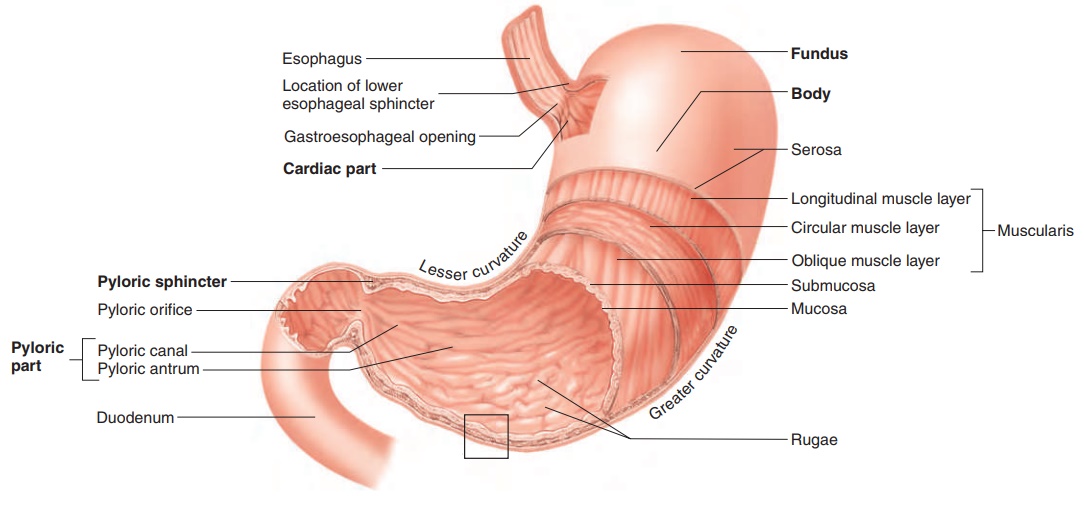
The stomach (figure 16.10) is an enlarged segment of the digestive tract in the left superior part of the abdomen. The opening from the esophagus into the stomach is called the gastroesophageal opening. The region of the stomach around the gastroesophageal opening is called the cardiac region because it is near the heart. The most supe-rior part of the stomach is the fundus (fŭn′ dŭs). The largest part of the stomach is the body, which turns to the right, forming a greatercurvature on the left and a lesser curvature on the right. The open-ing from the stomach into the small intestine is the pyloric (pı̄-lōr′ ik; gatekeeper) opening, which is surrounded by a relatively thick ring of smooth muscle called the pyloric sphincter. The region of the stomach near the pyloric opening is the pyloric region.
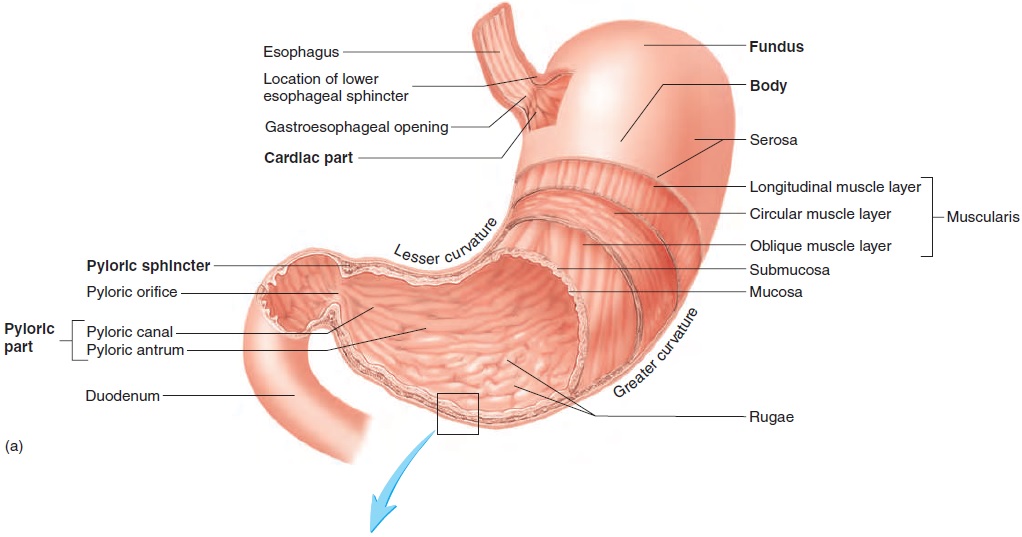
The muscular layer of the stomach is different from other regions of the digestive tract in that it consists of three layers: an outer longitudinal layer, a middle circular layer, and an inner oblique layer. These muscular layers produce a churning action in the stom-ach, important in the digestive process. The submucosa and mucosa of the stomach are thrown into large folds called rugae (roo′ gē; wrinkles) (figure 16.10a) when the stomach is empty. These folds allow the mucosa and submucosa to stretch, and the folds disappear as the stomach is filled.
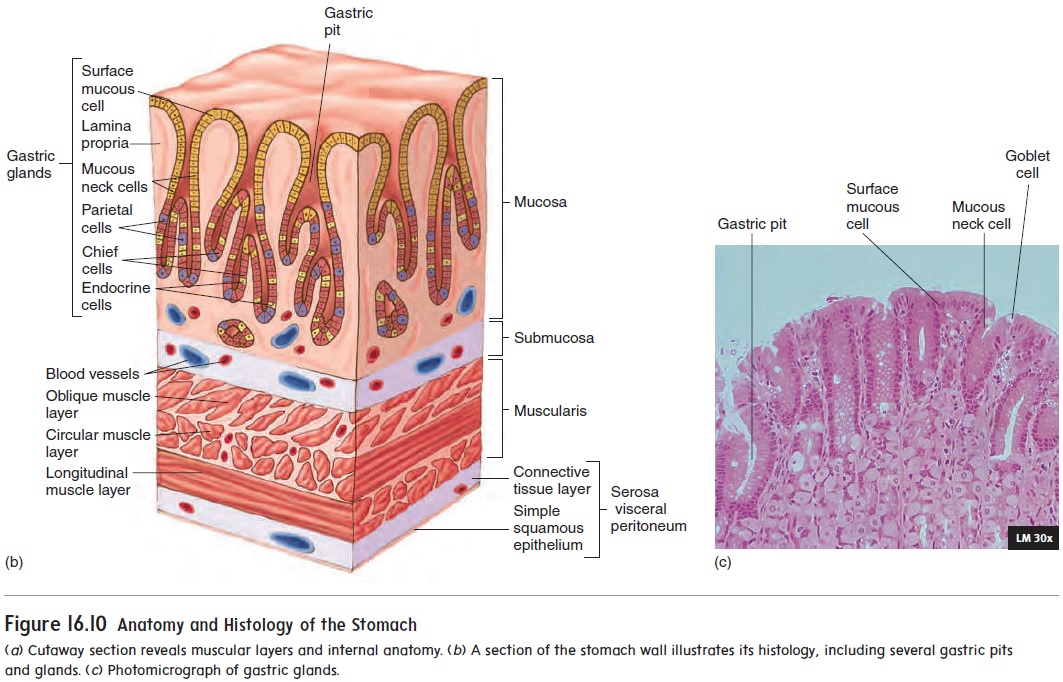
The stomach is lined with simple columnar epithelium. The mucosal surface forms numerous tubelike gastric pits (fig-ure 16.10b), which are the openings for the gastric glands. The epithelial cells of the stomach can be divided into five groups. The first group consists of surface mucous cells on the inner surface of the stomach and lining the gastric pits. Those cells produce mucus, which coats and protects the stomach lining. The remaining four cell types are in the gastric glands. They are
1. mucous neck cells, which produce mucus;
2. parietal cells, which produce hydrochloric acid andintrinsic factor;
3. endocrine cells, which produce regulatory chemicals; and
4. chief cells, which produce pepsinogen (pep-sin′ō-jen), aprecursor of the protein-digesting enzyme pepsin (pep′ sin; pepsis, digestion).
Secretions of the Stomach
The stomach functions primarily as a storage and mixing chamber for ingested food. As food enters the stomach, it is mixed with stomach secretions to become a semifluid mixture called chyme (kı̄m; juice). Although some digestion occurs in the stomach, that is not its principal function.
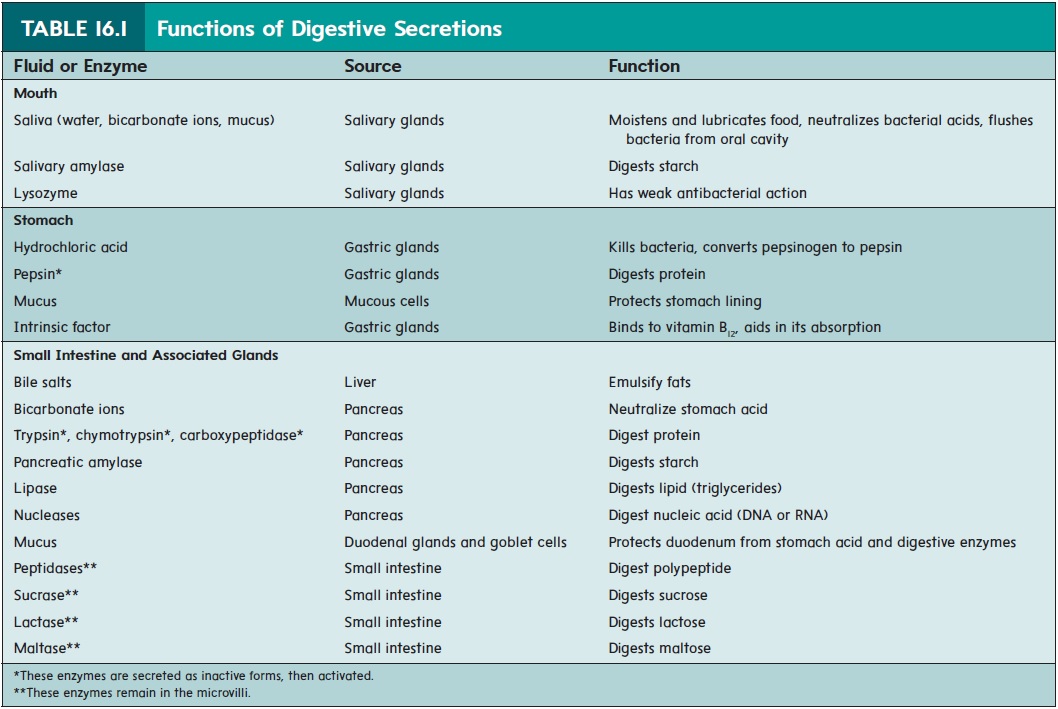
Stomach secretions from the gastric glands include hydrochloric acid, pepsin, mucus, and intrinsic factor (table 16.1). Hydrochloricacid produces a pH of about 2.0 in the stomach. The acid kills micro-organisms and activates pepsin from its inactive form, called pep-sinogen. Pepsin breaks covalent bonds of proteins to form smaller peptide chains. Pepsin exhibits optimum enzymatic activity at a pH of about 2.0. A thick layer of mucus lubricates the epithelial cells of the stomach wall and protects them from the damaging effect of the acidic chyme and pepsin. Irritation of the stomach mucosa stimulates the secretion of a greater volume of mucus. Intrinsic (in-trin′ sik) factor binds with vitamin B12 and makes it more readily absorbed in the small intestine. Vitamin B12 is important in deoxyri-bonucleic acid (DNA) synthesis and in red blood cell production.
Regulation of Stomach Secretions
Approximately 2 L of gastric secretions (gastric juice) are pro-duced each day. Both nervous and hormonal mechanisms regulate gastric secretions. The neural mechanisms involve central nervous system (CNS) reflexes integrated within the medulla oblongata. Higher brain centers can influence these reflexes. Local reflexes are integrated within the enteric plexus in the wall of the diges-tive tract and do not involve the CNS. Hormones produced by the stomach and intestine help regulate stomach secretions.
Regulation of stomach secretions can be divided into three phases: the cephalic, gastric, and intestinal phases. The cephalic phase can be viewed as the “get started” phase, when the stomach secretions are increased in anticipation of incoming food. This is followed by the gastric, “go for it,” phase, when most of the stimulation of secretion occurs. Finally, the intestinal phase is the “slow down” phase, during which stomach secretion decreases.
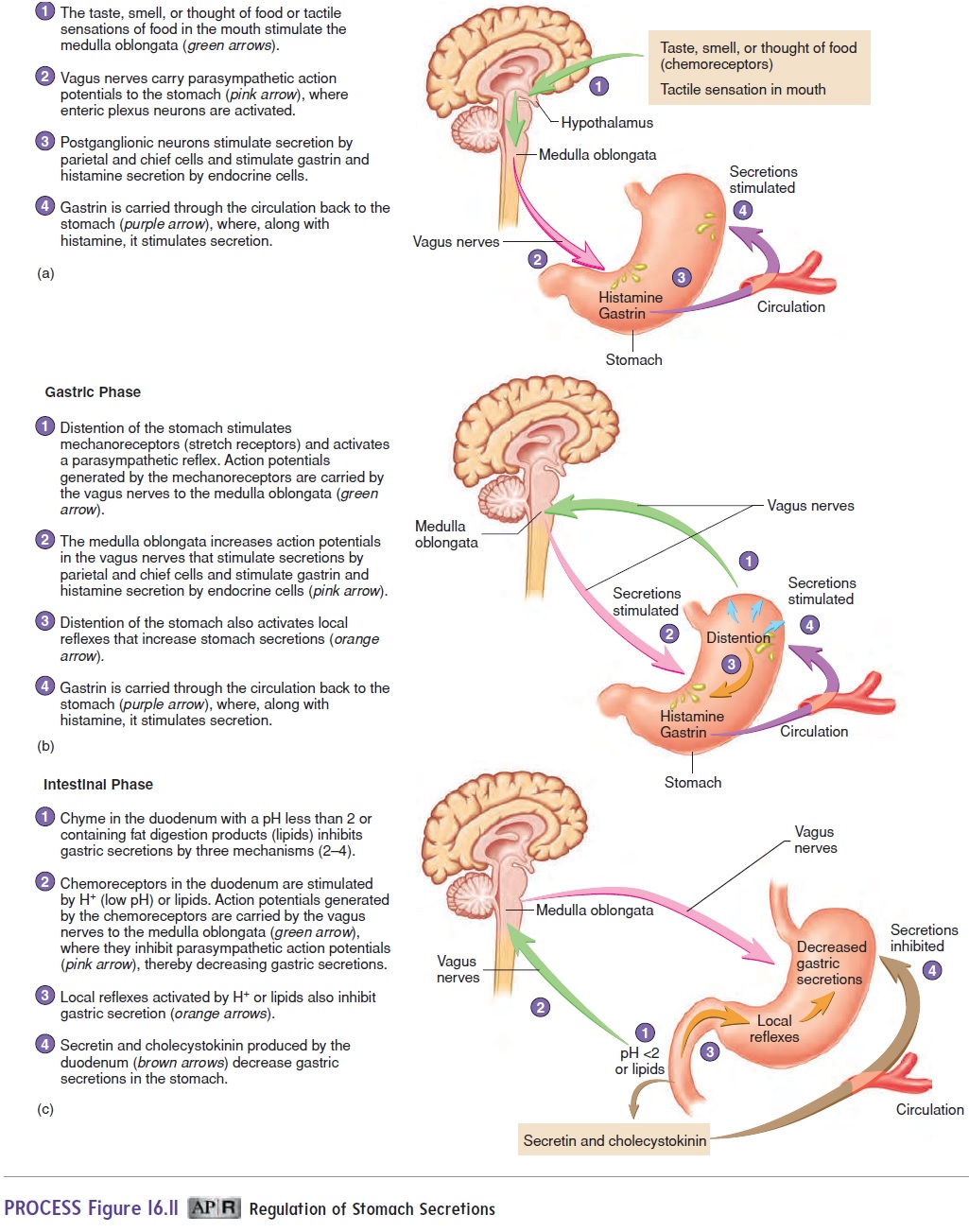
In the cephalic (se-fal′ ik; kephale, head) phase (figure 16.11a), sensations of taste, the smell of food, stimulation of tactile receptors during the process of chewing and swallowing, and pleasant thoughts of food stimulate centers within the medulla oblongata that influence gastric secretions. Action potentials are sent from the medulla oblongata along parasympathetic axons within the vagus nerves to the stomach. Within the stomach wall, the preganglionic neurons stimulate postganglionic neurons in the enteric plexus. The postganglionic neurons stimulate secretory activity in the cells of the stomach mucosa, causing the release of hydrochloric acid, pepsin, mucus, and intrinsic factor. The neurons also stimulate the release of gastrin and histamine from endocrine cells. Gastrin (gas′ trin) is a hormone that enters the circulation and is carried back to the stomach, where it stimulates additional secretory activity (table 16.2). Histamine is both a paracrine chem-ical signal that acts locally and a hormone that enters the blood to stimulate gastric gland secretory activity. Histamine is the most potent stimulator of hydrochloric acid secretion. Drugs that block the actions of histamine can lower acid levels.
The gastric phase is the period during which the greatest volume of gastric secretion occurs (figure 16.11b). The gastric phase is activated by the presence of food in the stomach. During the gastric phase, the food in the stomach is mixed with gastric secretions. Distention of the stomach stimulates stretch recep-tors. Action potentials generated by these receptors activate CNS reflexes and local reflexes, resulting in the cascade of events that increases secretion, as in the cephalic phase. Peptides, produced by the action of pepsin on proteins, stimulate the secretion of gastrin, which in turn stimulates additional hydrochloric acid secretion.
The intestinal phase of gastric secretion primarily inhibits gastric secretions (figure 16.11c). It is controlled by the entrance of acidic chyme into the duodenum, which initiates both neural and hormonal mechanisms. When the pH of the chyme enteringthe duodenum drops to 2.0 or below, the inhibitory influence of the intestinal phase is greatest. The hormone secretin (se-kr̄e ′ tin), which inhibits gastric secretions, is released from the duodenum in response to low pH (table 16.2). Fatty acids and peptides in the duo-denum initiate the release of the hormone cholecystokinin (k̄o ′ l̄e - sis-t̄o -k̄ı ′ nin), which also inhibits gastric secretions (table 16.2). Acidic chyme (pH < 2.0) in the duodenum also inhibits CNS stim-ulation and initiates local reflexes that inhibit gastric secretion.
To summarize, once gastric acid secretion begins, further secre-tion is controlled by negative-feedback loops involving nerves and hormones. First, during the gastric phase, high acid levels in the stomach trigger a decrease in additional acid secretion. Second, dur-ing the intestinal phase, acidic chyme entering the duodenum trig-gers a decrease in gastric acid secretion. These negative-feedback loops ensure that the acidic chyme entering the duodenum is neu-tralized, which is required for the digestion of food by pancreatic enzymes and for the prevention of peptic ulcer formation.
Movement In The Stomach
Two types of stomach movement aid digestion and help move chyme through the digestive tract: mixing waves and peristaltic waves (figure 16.12). Both types of movement result from smooth muscle contractions in the stomach wall. The contractions occur about every 20 seconds and proceed from the body of the stomach toward the pyloric sphincter. Relatively weak contractions result in mixingwaves,which thoroughly mix ingested food with stomach secretionsto form chyme. The more fluid part of the chyme is pushed toward the pyloric sphincter, whereas the more solid center moves back toward the body of the stomach. Stronger contractions result in peri-staltic waves, which force the chyme toward and through the pyloricsphincter. The pyloric sphincter usually remains closed because of mild tonic contraction. Each peristaltic contraction is sufficiently strong to cause partial relaxation of the pyloric sphincter and to pump a few milliliters of chyme through the pyloric opening and into the duodenum. Increased motility leads to increased emptying.
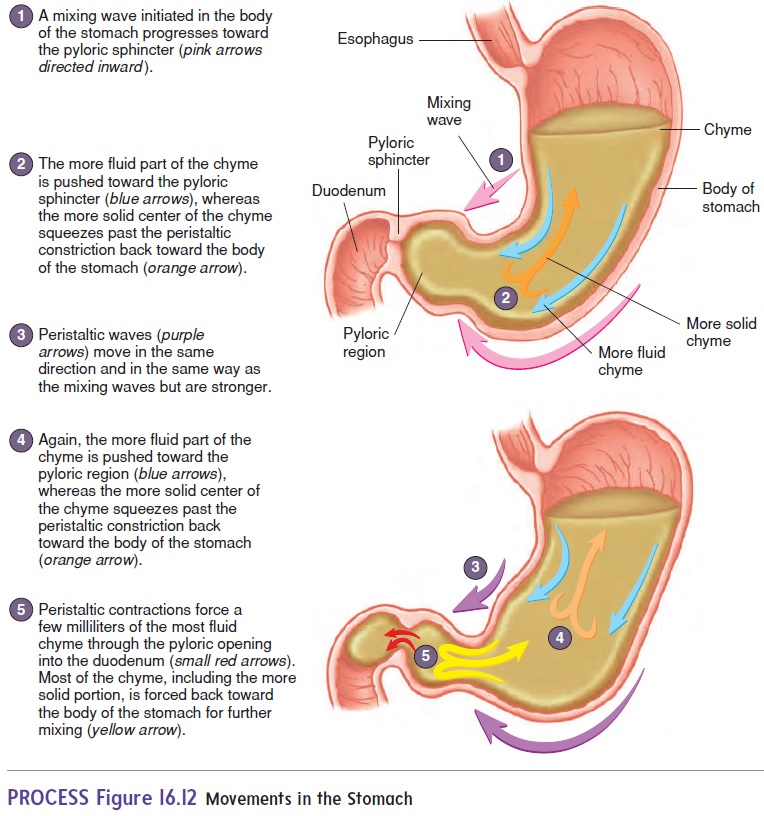
If the stomach empties too fast, the efficiency of digestion and absorption in the small intestine is reduced. However, if the rate of emptying is too slow, the highly acidic contents of the stomach may damage the stomach wall. To prevent these two extremes, stomach emptying is regulated. The hormonal and neural mecha-nisms that stimulate stomach secretions also increase stomach motility, so that the increased secretions are effectively mixed with the stomach contents. The major stimulus of gastric motility and emptying is distension of the stomach wall. Inhibition of gastric motility and emptying is accomplished by the same negative-feedback loops associated with the intestinal phase of gastric secretion. In particular, cholecystokinin is a major inhibitor of motility and emptying (table 16.2). Hence, stomach emptying is slower after a fatty meal due to the release of cholecystokinin.
In an empty stomach, peristaltic contractions that approach tetanic contractions can occur for about 2 to 3 minutes. The contractions are increased by low blood glucose levels and are sufficiently strong to create an uncomfortable sensation called a “hunger pang.” Hunger pangs usually begin 12 to 24 hours after the previous meal; in less time for some people. If nothing is ingested, hunger pangs reach their maximum intensity within 3 or 4 days and then become progressively weaker.
Related Topics