Chapter: Essentials of Anatomy and Physiology: Blood
Red Blood Cells: Function, Production and Maturation, Life Span, Blood Types
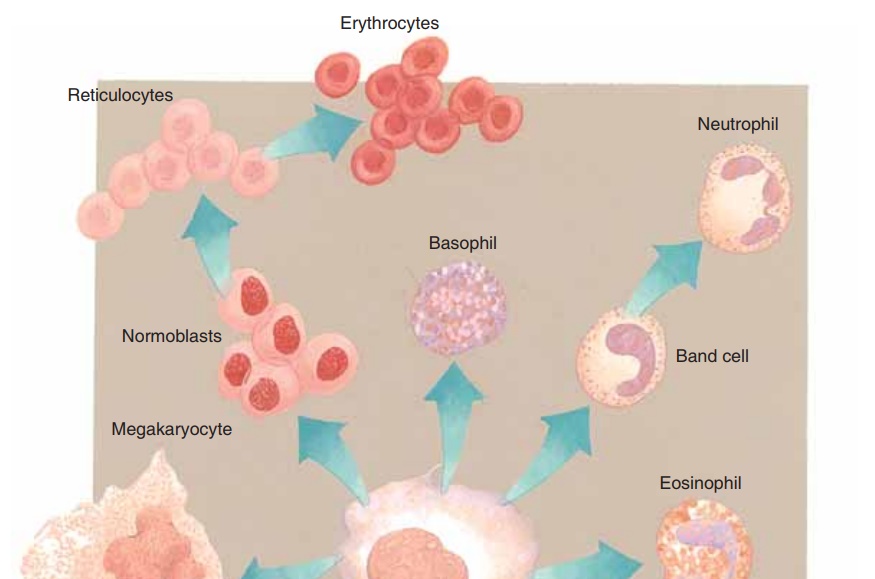
BLOOD CELLS
There are three kinds of blood cells: red blood cells, white blood cells, and platelets. Blood cells are pro-duced from stem cells inhemopoietic tissue. After birth this is primarily the red bone marrow, found in flat and irregular bones such as the sternum, hip bone, and vertebrae. Lymphocytes mature and divide in lymphatic tissue, found in the spleen, lymph nodes, and thymus gland. The thymus contains stem cells that produce T lymphocytes, and the stem cells in other lymphatic tissue also produce lympho-cytes.
RED BLOOD CELLS
Also called erythrocytes, red blood cells (RBCs) are biconcave discs, which means their centers are thinner than their edges. You may recall that red blood cells are the only human cells with-out nuclei. Their nuclei disintegrate as the red blood cells mature and are not needed for normal func-tioning.
A normal RBC count ranges from 4.5 to 6.0 million cells per microliter (mL) of blood (1 microliter 5 1 mm3 5 one millionth of a liter, a very small volume). RBC counts for men are often toward the high end of this range; those for women are often toward the low end. Another way to measure the amount of RBCs is the hematocrit. This test involves drawing blood into a thin glass tube called a capillary tube, and centrifug-ing the tube to force all the cells to one end. The per-centages of cells and plasma can then be determined. Because RBCs are by far the most abundant of the blood cells, a normal hematocrit range is just like that of the total blood cells: 38% to 48%. Both RBC count and hematocrit (Hct) are part of a complete blood count (CBC).
Function
Red blood cells contain the protein hemoglobin (Hb), which gives them the ability to carry oxygen. Each red blood cell contains approximately 300 mil-lion hemoglobin molecules, each of which can bond to four oxygen molecules. In the pul-monary capillaries, RBCs pick up oxygen and oxyhe-moglobin is formed. In the systemic capillaries, hemoglobin gives up much of its oxygen and becomes reduced hemoglobin. A determination of hemoglobin level is also part of a CBC; the normal range is 12 to 18 grams per 100 mL of blood. Essential to the for-mation of hemoglobin is the mineral iron; there are four atoms of iron in each molecule of hemoglobin. It is the iron that actually bonds to the oxygen and also makes RBCs red.
Hemoglobin is also able to bond to carbon dioxide (CO2), and does transport some CO2 from the tissues to the lungs. But hemoglobin accounts for only about 10% of total CO2 transport (most is carried in the plasma as bicarbonate ions).
Production and Maturation
Red blood cells are formed in red bone marrow (RBM) in flat and irregular bones. Within red bone marrow are precursor cells calledstem cells. Recall that stem cells are unspecialized cells that may develop, or differentiate, in several ways. The stem cells of the red bone marrow may also be called hemocytoblasts (hemo 5 “blood,” cyto 5 “cell,” blast 5 “producing”), and they constantly undergo mitosis to produce all the kinds of blood cells, many of which are RBCs (Figs. 11–2 and 11–3). The rate of produc-tion is very rapid (estimated at several million new RBCs per second), and a major regulating factor is oxygen. If the body is in a state of hypoxia, or lack of oxygen, the kidneys produce a hormone called ery-thropoietin, which stimulates the red bone marrow to increase the rate of RBC production (that is, the rate of stem cell mitosis). This will occur following hemor-rhage or if a person stays for a time at a higher altitude. As a result of the action of erythropoietin, more RBCs will be available to carry oxygen and correct the hypoxic state.
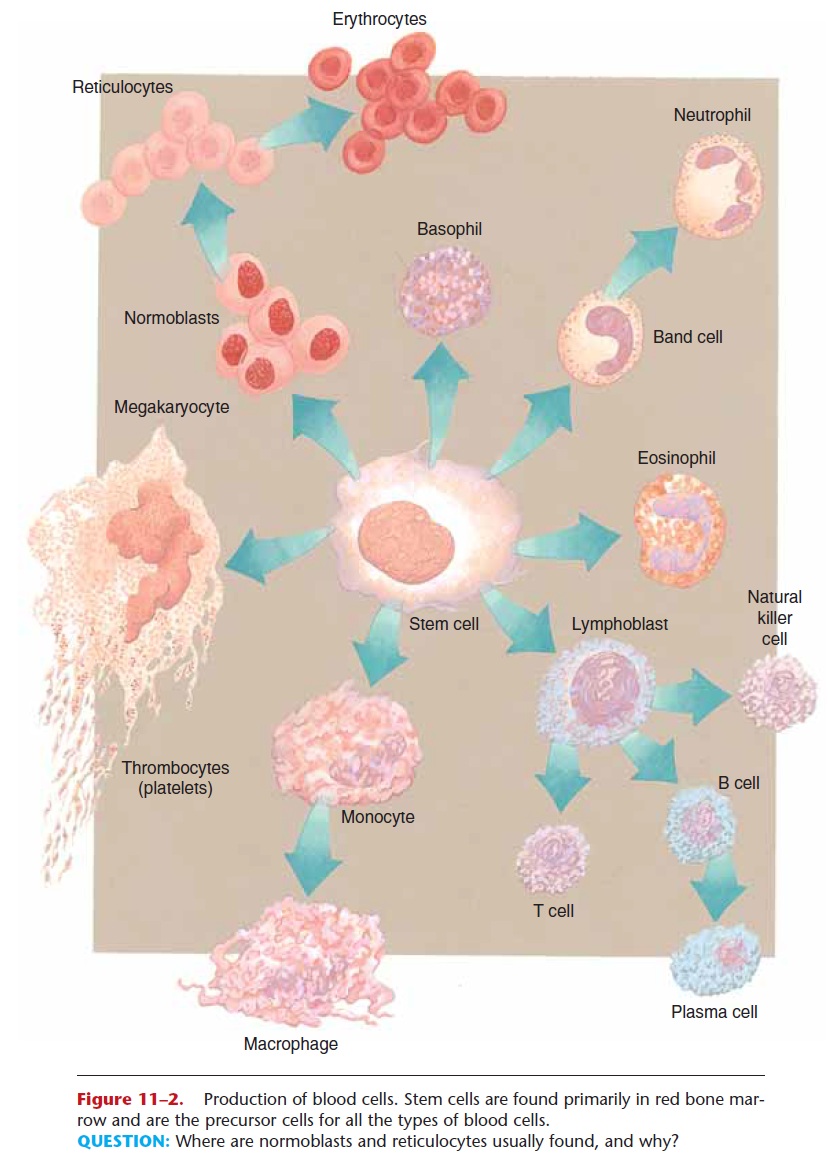
Figure 11–2. Production of blood cells. Stem cells are found primarily in red bone mar-row and are the precursor cells for all the types of blood cells.
QUESTION: Where are normoblasts and reticulocytes usually found, and why?
The stem cells that will become RBCs go through a number of developmental stages, only the last two of which we will mention (see Fig. 11–2). The nor-moblast is the last stage with a nucleus, which then disintegrates. The reticulocyte has fragments of the endoplasmic reticulum, which are visible when blood smears are stained for microscopic evaluation. These immature cells are usually found in the red bone mar-row, although a small number of reticulocytes in the peripheral circulation is considered normal (up to 1.5% of the total RBCs). Large numbers of reticulo-cytes or normoblasts in the circulating blood mean that the number of mature RBCs is not sufficient to carry the oxygen needed by the body. Such situations include hemorrhage, or when mature RBCs have been destroyed, as in Rh disease of the newborn, and malaria.
The maturation of red blood cells requires many nutrients. Protein and iron are necessary for the syn-thesis of hemoglobin and become part of hemoglobin molecules. Copper is part of some enzymes involved in hemoglobin synthesis. The vitamins folic acid and B12 are required for DNA synthesis in the stem cells of the red bone marrow. As these cells undergo mitosis, they must continually produce new sets of chromo-somes. Vitamin B12 is also called the extrinsic factor because its source is external, our food. Parietal cells of the stomach lining produce the intrinsic factor, a chemical that combines with the vitamin B12 in food to prevent its digestion and promote its absorption in the small intestine. A deficiency of either vitamin B12 or the intrinsic factor results in pernicious anemia.
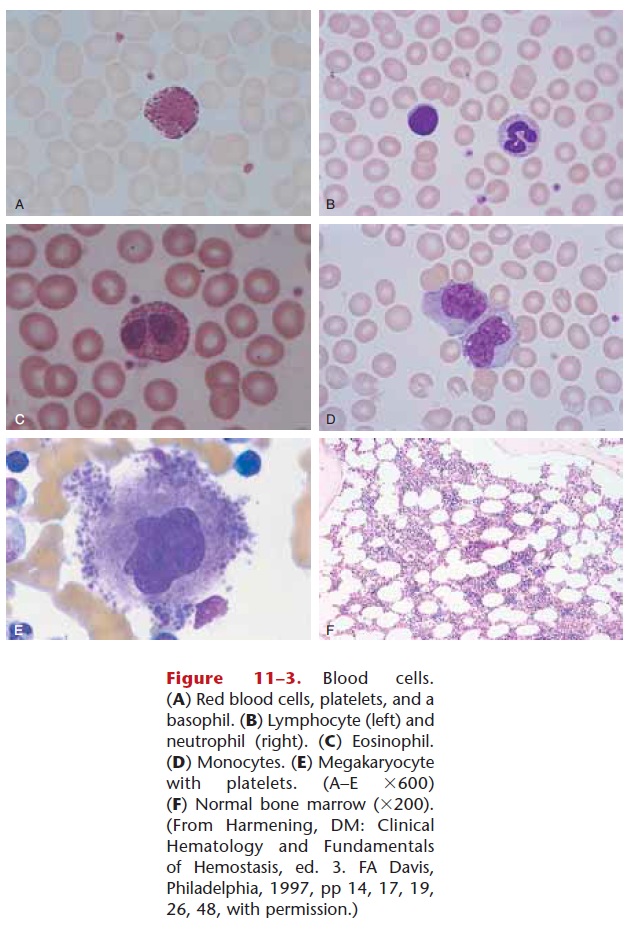
Life Span
Red blood cells live for approximately 120 days. As they reach this age they become fragile and are removed from circulation by cells of the tissue macrophage system (formerly called the reticuloen-dothelial or RE system). The organs that contain macrophages (literally, “big eaters”) are the liver, spleen, and red bone marrow. Look at Fig. 11–4 as you read the following. The old RBCs are phagocytized and digested by macrophages, and the iron they con-tained is put into the blood to be returned to the red bone marrow to be used for the synthesis of new hemoglobin. If not needed immediately for this pur-pose, excess iron is stored in the liver. The iron of
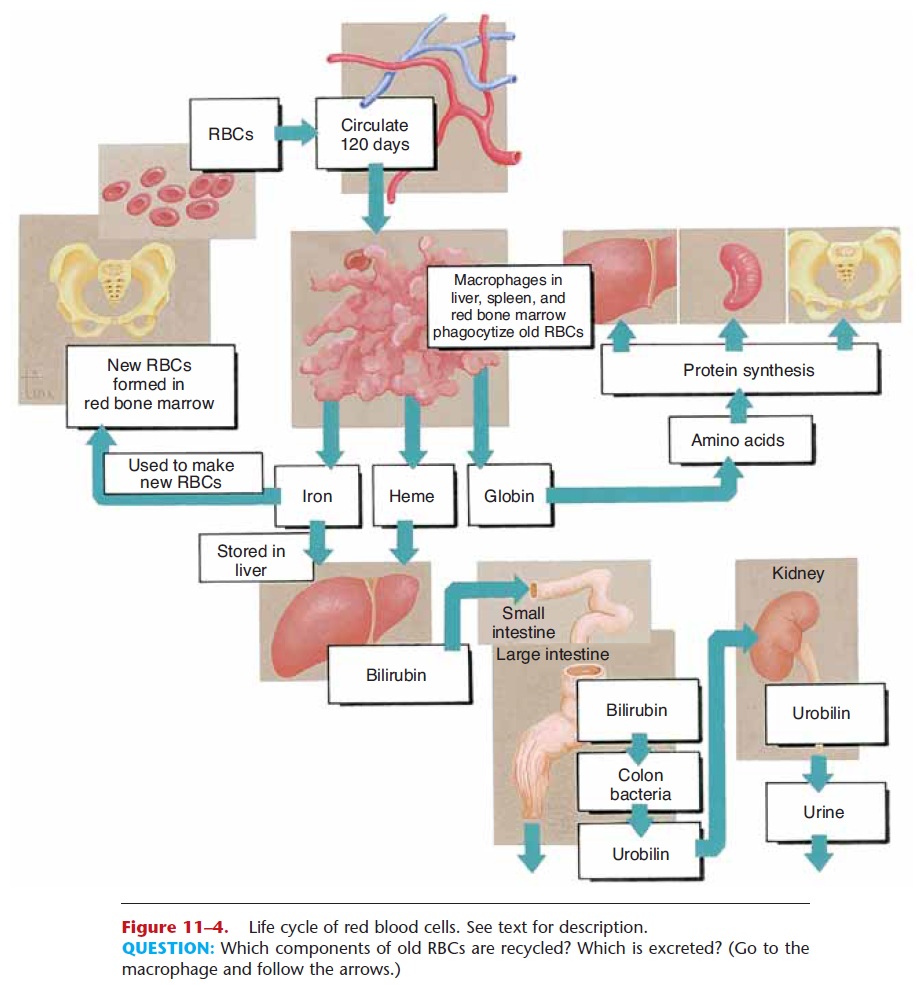
Figure 11–4. Life cycle of red blood cells. See text for description.
QUESTION: Which components of old RBCs are recycled? Which is excreted? (Go to the macrophage and follow the arrows.)
RBCs is actually recycled over and over again. The globin or protein portion of the hemoglobin molecule is also recycled. It is digested to its amino acids, which may then be used for the synthesis of new proteins.
Another part of the hemoglobin molecule is the heme portion, which cannot be recycled and is a waste product. The heme is converted to bilirubin by macrophages. The liver removes bilirubin from circu-lation and excretes it into bile; bilirubin is a bile pig-ment. Bile is secreted by the liver into the duodenum and passes through the small intestine and colon, so bilirubin is eliminated in feces, and gives feces their characteristic brown color. In the colon some bilirubin is changed to urobilinogen by the colon bacteria.
Some urobilinogen may be absorbed into the blood, but it is changed to urobilin and excreted by the kidneys in urine. If bilirubin is not excreted properly, perhaps because of liver disease such as hepatitis, it remains in the blood. This may cause jaundice, a condition in which the whites of the eyes appear yellow. This yellow color may also be seen in the skin of lightskinned people.
Blood Types
Our blood types are genetic; that is, we inherit genes from our parents that determine our own types. There are many red blood cell factors or types; we will discuss the two most important ones: the ABO group and the Rh factor.
The ABO group contains four blood types: A, B, AB, and O. The letters A and B represent antigens (protein-oligosaccharides) on the red blood cell mem-brane. A person with type A blood has the A antigen on the RBCs, and someone with type B blood has the B antigen. Type AB means that both A and B antigens are present, and type O means that neither the A nor the B antigen is present.
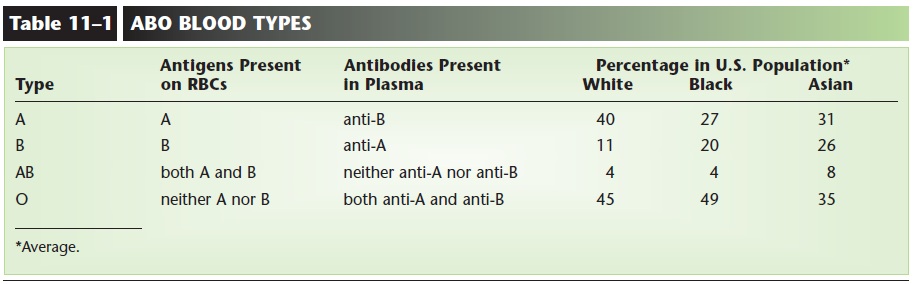
Circulating in the plasma of each person are natu-ral antibodies for those antigens not present on the RBCs. Therefore, a type A person has anti-B antibod-ies in the plasma; a type B person has anti-A antibod-ies; a type AB person has neither anti-A nor anti-B antibodies; and a type O person has both anti-A and anti-B antibodies (see Table 11–1 and Fig. 11–5).
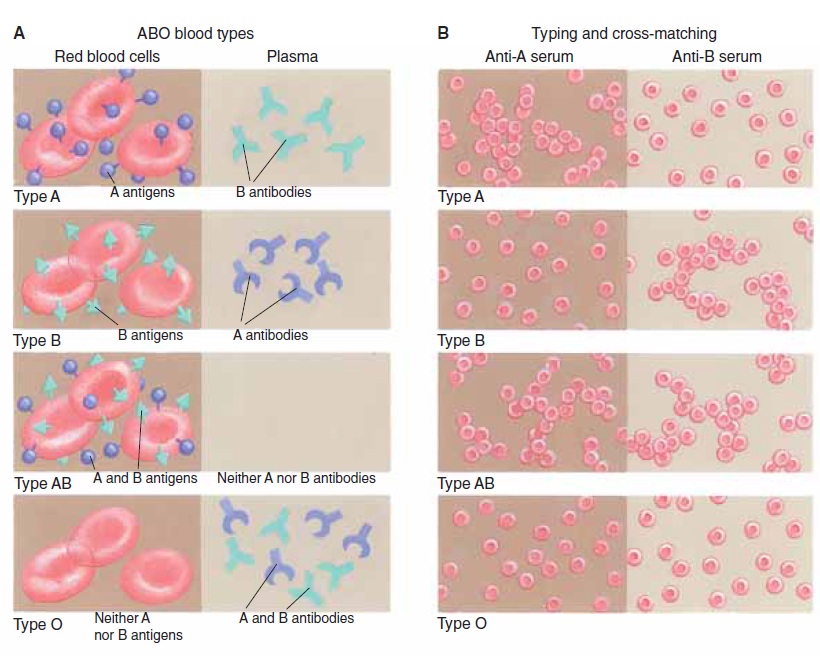
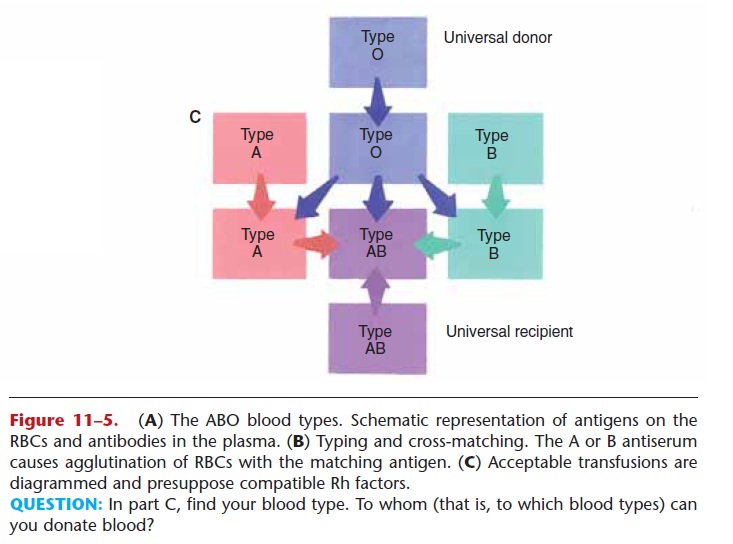
Figure 11–5. (A) The ABO blood types. Schematic representation of antigens on the RBCs and antibodies in the plasma. (B) Typing and cross-matching. The A or B antiserum causes agglutination of RBCs with the matching antigen. (C) Acceptable transfusions are diagrammed and presuppose compatible Rh factors.
QUESTION: In part C, find your blood type. To whom (that is, to which blood types) can you donate blood?
These natural antibodies are of great importance for transfusions. If possible, a person should receive blood of his or her own type; only if this type is not available should another type be given. For example, let us say that a type A person needs a transfusion to replace blood lost in hemorrhage. If this person were to receive type B blood, what would happen? The type A recipient has anti-B antibodies that would bind to the type B antigens of the RBCs of the donated blood. The type B RBCs would first clump (agglutination) then rupture (hemolysis), thus defeating the purpose of the transfusion. An even more serious consequence is that the hemoglobin of the ruptured RBCs, now called free hemoglobin, may clog the capillaries of the kidneys and lead to renal damage or renal failure. You can see why typing and cross-matching of donor and recipient blood in the hospital laboratory is so impor-tant before any transfusion is given (see Fig. 11–5). This procedure helps ensure that donated blood will not bring about a hemolytic transfusion reaction in the recipient.
You may have heard of the concept that a person with type O blood is a “universal donor.” Usually, a unit of type O negative blood may be given to people with any other blood type. This is so because type O RBCs have neither the A nor the B antigens and will not react with whatever antibodies the recipient may have. If only one unit (1 pint) of blood is given, the anti-A and anti-B antibodies in the type O blood plasma will be diluted in the recipient’s blood plasma and will not have a harmful effect on the recipient’s RBCs. The term negative, in O negative, the universal donor, refers to the Rh factor, which we will now consider.
The Rh factor is another antigen (often called D) that may be present on RBCs. People whose RBCs have the Rh antigen are Rh positive; those without the antigen are Rh negative. Rh-negative people do not have natural antibodies to the Rh antigen, and for them this antigen is foreign. If an Rh-negative person receives Rh-positive blood by mistake, antibodies will be formed just as they would be to bacteria or viruses. A first mistaken transfusion often does not cause problems, because antibody pro-duction is slow upon the first exposure to Rh-positive RBCs. A second transfusion, however, when anti-Rh antibodies are already present, will bring about a transfusion reaction, with hemolysis and possible kid-ney damage.
Related Topics