Chapter: Essentials of Anatomy and Physiology: Blood
Formed Elements - Blood
Formed Elements
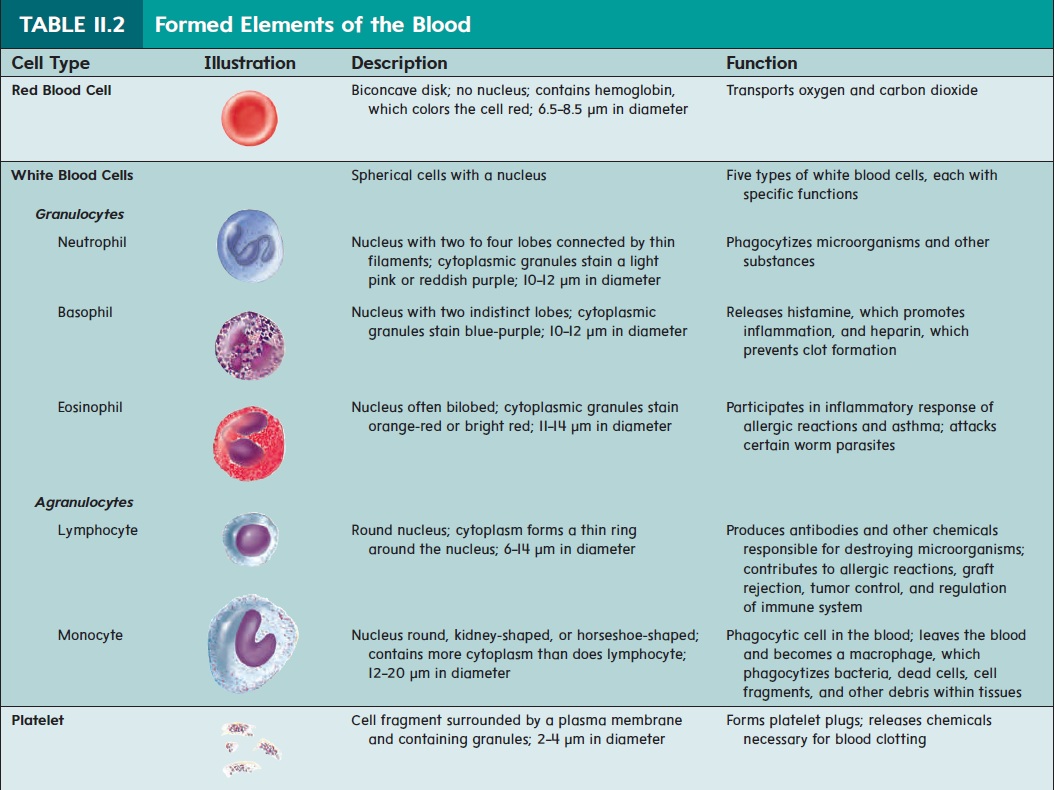
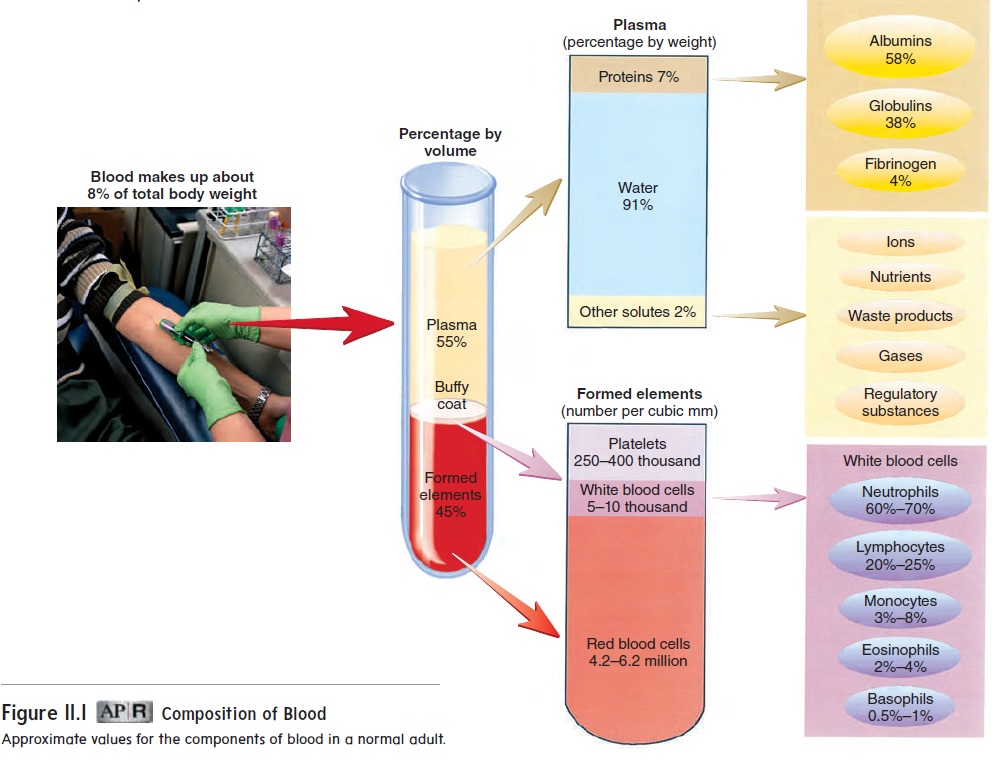
About 95% of the volume of the formed elements consists of redblood cells (RBCs), orerythrocytes(ĕ-rith′rō-sı̄tz;erythro-,red+ kytos, cell). The remaining 5% of the volume of the formed elements consists ofwhite blood cells (WBCs), or leukocytes(loo′kō-sı̄tz; leuko-, white), and cell fragments called platelets (plāt′letz), or thrombocytes (throm′bō-sı̄tz; thrombo-, clot). Redblood cells are 700 times more numerous than white blood cells and 17 times more numerous than platelets. Table 11.2 illustrates the formed elements of the blood.
Production of Formed Elements
The process of blood cell production is called hematopoiesis (hē′mă-tō-poy-ē′sis; hemato-, blood + poiesis, making). In the fetus, hematopoiesis occurs in several tissues, including the liver, thymus, spleen, lymph nodes, and red bone marrow. After birth, hematopoiesis is confined primarily to red bone marrow, but some white blood cells are produced in lymphatic tissues .
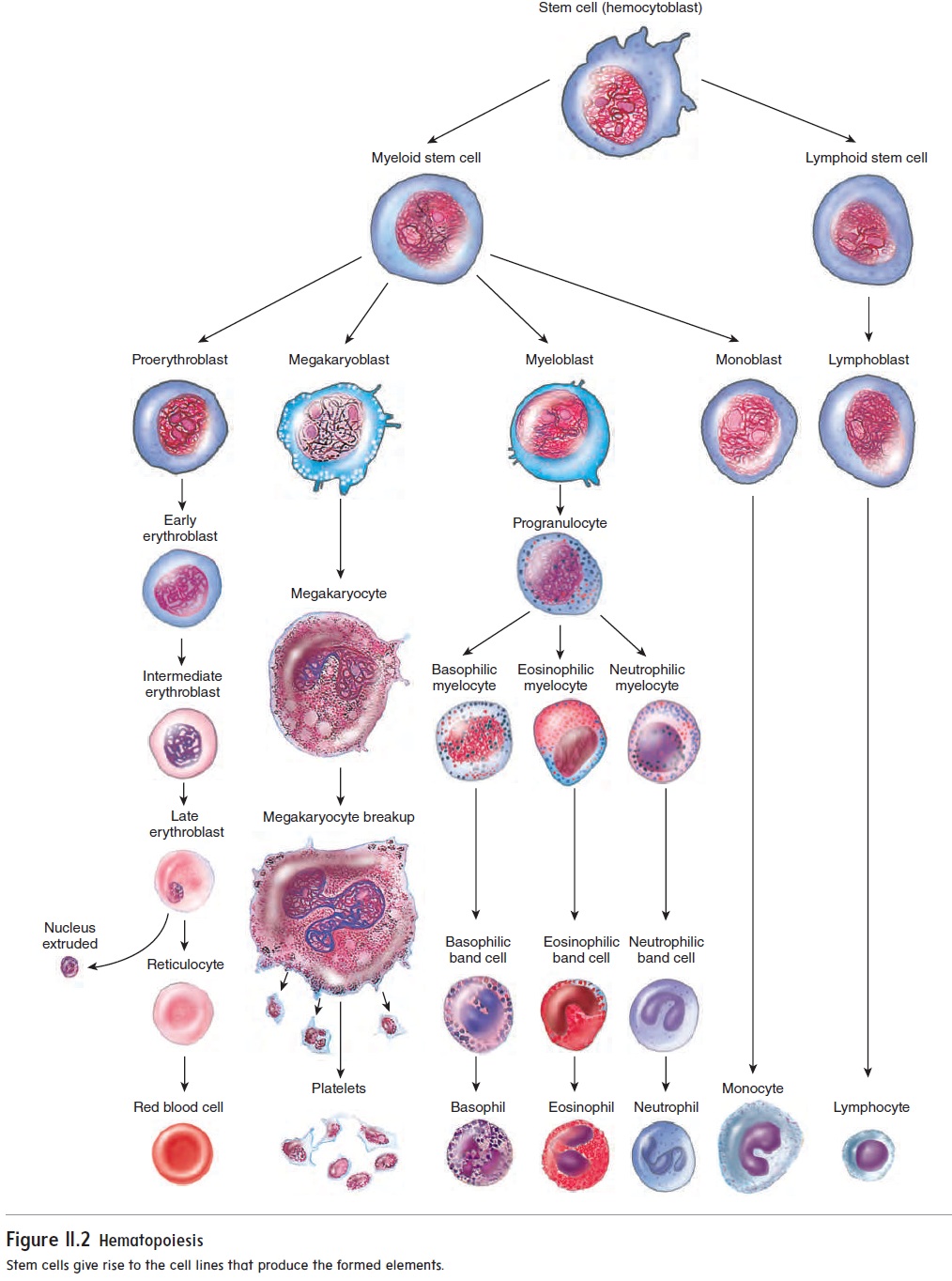
All the formed elements of blood are derived from a single population of cells called stem cells, or hemocytoblasts. These stem cells differentiate to give rise to different cell lines, each of which ends with the formation of a particular type of formed ele-ment (figure 11.2). The development of each cell line is regulated by specific growth factors. That is, growth factors determine the types of formed elements derived from the stem cells and how many formed elements are produced.
Red Blood Cells
Normal red blood cells are disk-shaped, with edges that are thicker than the center of the cell (figure 11.3). The biconcave shape increases the cell’s surface area compared to a flat disk of the same size. The greater surface area makes it easier for gases to move into and out of the red blood cell. In addition, the red blood cell can bend or fold around its thin center, decreasing its size and enabling it to pass more easily through smaller blood vessels.
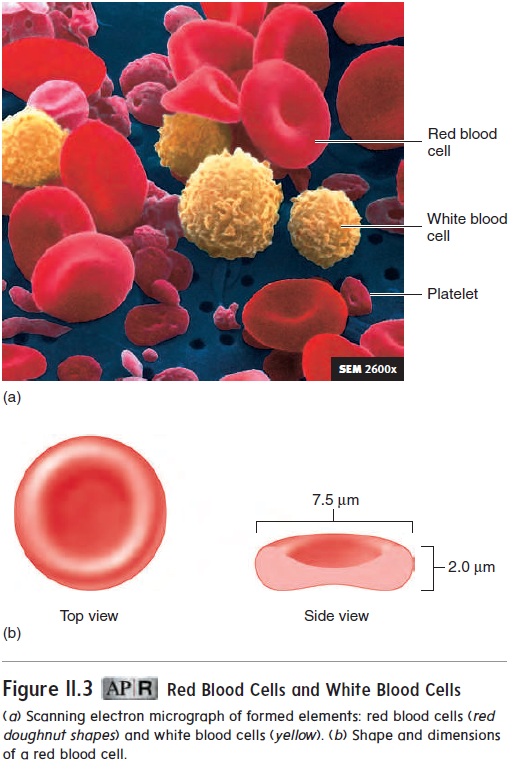
During their development, red blood cells lose their nuclei and most of their organelles. Consequently, they are unable to divide. Red blood cells live for about 120 days in males and 110 days in females. One-third of a red blood cell’s volume is the pigmented protein hemoglobin (hē-mō-glō′bin), which is responsible for the cell’s red color.
Function
The primary functions of red blood cells are to transport oxygen from the lungs to the various tissues of the body and to help transport carbon dioxide from the tissues to the lungs. Oxygen transport is accomplished by hemoglobin, which consists of four protein chains and four heme groups. Each protein, called a globin (gl̄o ′ bin), is bound to one heme (h̄e m), a red-pigmented molecule.
Each heme contains one iron atom, which is necessary for the normal function of hemoglobin. Each iron in a heme molecule can reversibly bind to an oxygen molecule. Hemoglobin picks up oxygen in the lungs and releases oxygen in other tissues (see chap-ter 15). Hemoglobin that is bound to oxygen is bright red, whereashemoglobin without bound oxygen is a darker red. Hemoglobin is responsible for 98.5% of the oxygen transported in blood. The remaining 1.5% is transported dissolved in plasma.
Because iron is necessary for oxygen transport, it is not surpris-ing that two-thirds of the body’s iron is found in hemoglobin. Small amounts of iron are required in the diet to replace the small amounts lost in the urine and feces, but otherwise the existing iron is recycled, as described later in this section. Women need more dietary iron than men do because women lose iron as a result of menstruation.
Although oxygen is the primary molecule that binds to hemoglobin, other molecules can also bind to hemoglobin. Carbon monoxide, a gas produced by the incomplete combustion of hydrocarbons, such as gasoline, is one example. It binds to the iron in hemoglobin about 210 times more readily than does oxygen and does not tend to unbind. As a result, the hemoglobin bound to carbon monoxide no longer transports oxygen. Nausea, headache, unconsciousness, and death are possible consequences of prolonged exposure to carbon monoxide.
Carbon dioxide is produced in tissues and transported in the blood to the lungs, where it is removed from the blood . Carbon dioxide transport involves bicarbonate ions, hemoglobin, and plasma. Approximately 70% of the carbon dioxide in blood is transported in the form of bicarbonate ions. The enzyme carbonic anhydrase (kar-bon′ ik an-hı̄′ drās), found primarily inside red blood cells, catalyzes a reaction that converts carbon dioxide (CO2) and water (H2O) into a hydrogen ion (H+) and a bicarbonate ion (HCO3−):
CO2 + H2O < -- -- > H+ + HCO3−
Carbon dioxide can bind reversibly to the globin part of hemoglobin. About 23% of the CO2 in blood is transported bound to hemoglobin or other blood proteins. The remaining 7% of CO2 is transported dissolved in plasma.
Life History of Red Blood Cells
Under normal conditions, about 2.5 million red blood cells are destroyed every second. Fortunately, new red blood cells are pro-duced just as rapidly. Stem cells form proerythroblasts (prō-ĕ-rith′ rō-blastz;pro-, before + erythro-, red + blastos, germ), which give rise to the red blood cell line (see figure 11.2). Red blood cell production involves a series of cell divisions. After each cell divi-sion, the new cells change and become more like mature red blood cells. In the later divisions, the newly formed cells manufacture large amounts of hemoglobin. After the final cell division, the cells lose their nuclei and become completely mature red blood cells.
The process of cell division requires the B vitamins folate and B12, which are necessary for the synthesis of DNA . Iron is required for the production of hemoglobin. Consequently, a lack of folate, vitamin B12, or iron can interfere with normal red blood cell production.
Red blood cell production is stimulated by low blood oxygen levels. Typical causes of low blood oxygen are decreased numbers of red blood cells, decreased or defective hemoglobin, diseases of the lungs, high altitude, inability of the cardiovascular system to deliver blood to tissues, and increased tissue demand for oxygen, as occurs during endurance exercises.
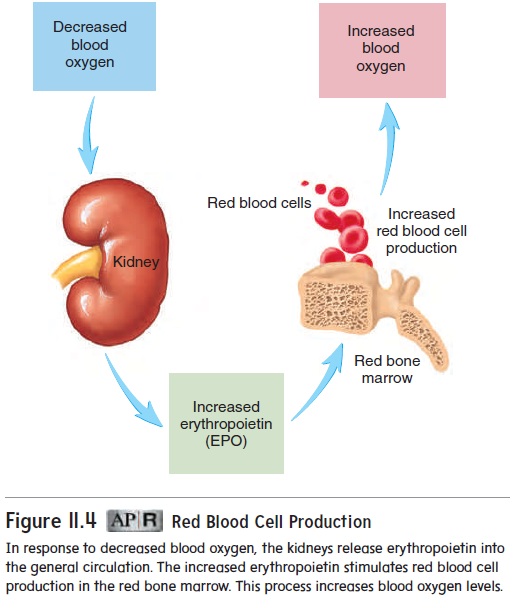
Low blood oxygen levels stimulate red blood cell produc-tion by increasing the formation and release of the glycoprotein erythropoietin (ĕ-rith-rō-poy′ĕ-tin, EPO), primarily by the kid-neys (figure 11.4). Erythropoietin stimulates red bone marrow to produce more red blood cells. Thus, when oxygen levels in the blood decrease, the production of erythropoietin increases, which increases red blood cell production. The greater number of red blood cells increases the blood’s ability to transport oxygen. This negative-feedback mechanism increases the blood’s capacity to transport oxygen and maintains homeostasis. Conversely, if blood oxygen levels rise, less erythropoietin is released, and red blood cell production decreases.
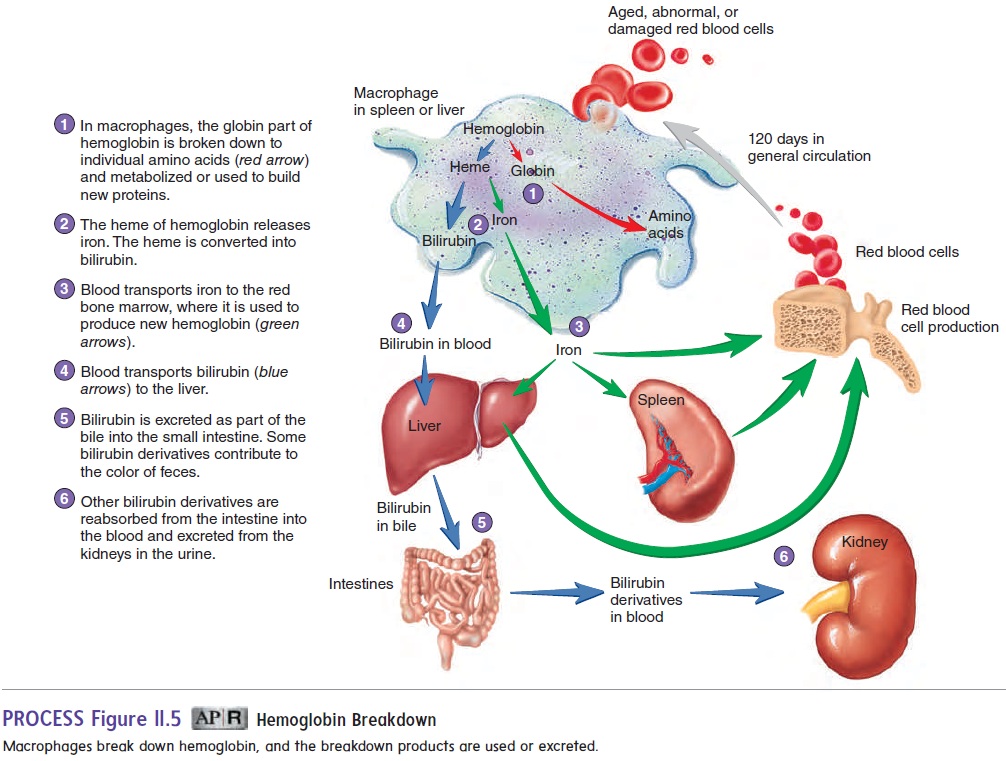
When red blood cells become old, abnormal, or damaged, they are removed from the blood by macrophages located in the spleen and liver (figure 11.5). Within the macrophage, the globin part of the hemoglobin molecule is broken down into amino acids that are reused to produce other proteins. The iron released from heme is transported in the blood to the red bone marrow and used to produce new hemoglobin. Thus, the iron is recycled. The heme molecules are converted to bilirubin (bil-i-roo′ bin), a yellow pig-ment molecule. Bilirubin is normally taken up by the liver and released into the small intestine as part of the bile . If the liver is not functioning normally, or if the flow of bile from the liver to the small intestine is hindered, bilirubin builds up in the circulation and produces jaundice (jawn′ dis;jaune, yellow), a yellowish color to the skin. After it enters the intestine, bilirubin is converted by bacteria into other pigments.
Some of these pigments give feces their brown color, whereas others are absorbed from the intestine into the blood, modified by the kidneys, and excreted in the urine, contributing to the characteristic yellow color of urine.
White Blood Cells
White blood cells are spherical cells that lack hemoglobin. When the components of blood are separated from one another, whiteblood cells as well as platelets make up the buffy coat, a thin, white layer of cells between plasma and red blood cells (see figure 11.1). White blood cells are larger than red blood cells, and each has a nucleus (table 11.2). Although white blood cells are components of the blood, the blood serves primarily as a means of transporting these cells to other body tissues. White blood cells can leave the blood and travel by ameboid (ă-mē′ boyd; like an ameba) movement through the tissues. In this process, the cell projects a cytoplasmic extension that attaches to an object. Then the rest of the cell’s cyto-plasm flows into the extension. Two functions of white blood cells are (1) to protect the body against invading microorganisms and other pathogens and (2) to remove dead cells and debris from the tissues by phagocytosis.
Each white blood cell type is named according to its appear-ance in stained preparations. Those containing large cytoplasmic granules are granulocytes (gran′ ū-lō-sı̄tz; granulo-, granular), and those with very small granules that cannot be seen easily with the light microscope are agranulocytes (ă-gran′ ū-lō-sı̄tz; a-, without).
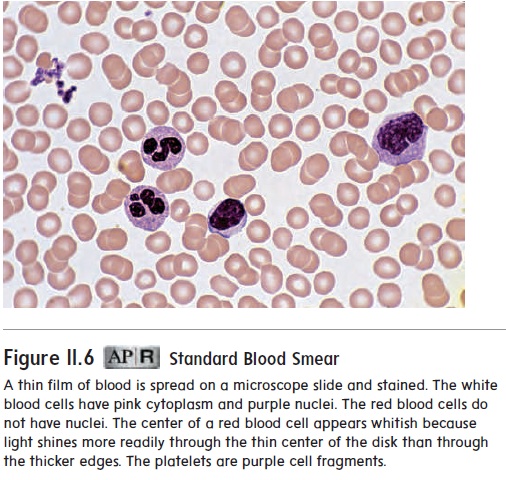
There are three kinds of granulocytes: neutrophils, basophils, and eosinophils. Neutrophils (noo′ trō-filz; neutro-, neutral + philos, loving), the most common type of white blood cells, havesmall cytoplasmic granules that stain with both acidic and basic dyes (figure 11.6). Their nuclei are commonly lobed, with the number of lobes varying from two to four. Neutrophils usually remain in the blood for a short time (10–12 hours), move into other tissues, and phagocytize microorganisms and other foreign substances. Dead neutrophils, cell debris, and fluid can accumu-late as pus at sites of infections.
Basophils (bā′sō-filz;baso-,base), the least common of allwhite blood cells, contain large cytoplasmic granules that stain blue or purple with basic dyes (table 11.2). Basophils release histamine and other chemicals that promote inflammation. They also release heparin, which prevents the formation of clots.
Eosinophils (ē-ō-sin′o-filz) contain cytoplasmic granules thatstain bright red with eosin, an acidic stain. They often have a two-lobed nucleus (table 11.2). Eosinophils are involved in inflammatory responses associated with allergies and asthma. In addition, chemicals from eosinophils are involved in destroying certain worm parasites.
There are two kinds of agranulocytes: lymphocytes and mono-cytes. Lymphocytes (lim′ fō-sı̄tz; lympho-, lymph) are the smallest of the white blood cells (table 11.2). The lymphocytic cytoplasm consists of only a thin, sometimes imperceptible ring around the nucleus. There are several types of lymphocytes, and they play an important role in the body’s immune response. Their diverse activities involve the production of antibodies and other chemicals that destroy microorganisms, contribute to allergic reactions, reject grafts, control tumors, and regulate the immune system.
Monocytes (mon′ō-sı̄tz) are the largest of the white bloodcells (table 11.2). After they leave the blood and enter tissues, monocytes enlarge and become macrophages (mak′ rō-fā-jez; macro-, large+ phagō, to eat), which phagocytize bacteria, deadcells, cell fragments, and any other debris within the tissues. In addition, macrophages can break down phagocytized foreign substances and present the processed substances to lymphocytes, causing activation of the lymphocytes .
Platelets
Platelets are minute fragments of cells, each consisting of a small amount of cytoplasm surrounded by a cell membrane (see figure 11.6). They are produced in the red bone marrow from megakaryocytes(meg-ă-kar′ē-ō-sı̄tz;mega-,large+karyon,nucleus), which are large cells (see figure 11.2). Small fragments of these cells break off and enter the blood as platelets, which play an important role in preventing blood loss.
Related Topics