Chapter: Genetics and Molecular Biology: Genetics
Principles of Bacterial Sex
Principles of Bacterial Sex
Lederberg and Tatum discovered that some bacterial
strains can trans-fer DNA to recipient cells, that is, they can mate. This
discovery opened the doors for two types of research. First, genetic
manipulations could be used to assist other types of studies involving bacteria,
and through-out the book we shall see many examples of the assistance genetics
provides to biochemical, physiological, and physical studies. Second, the
mechanism of bacterial mating itself was interesting and could be investigated.
In this section we shall review the actual mechanism of bacterial mating.
Maleness is conferred upon cells when they contain
a mating module called the F-factor. F stands for fertility. This module is a
circular DNA molecule containing about 25 genes involved with conjugation. One
of the F genes codes for the major protein of the F-pilus. This is an appendage
essential for DNA transfer. Other genes code for additional parts of the
F-pilus, its membrane attachment, and DNA replication of F-factor DNA. The
system also contains at least one regulatory gene.
Figure 8.19 Transfer of one strand of the
double-stranded F factor from malecells into female cells.
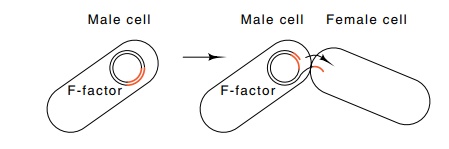
F-pilus
contact with a suitable female activates the mating module. As a result, a
break is made within the F sequences of DNA and a single strand of DNA
synthesized via the rolling circle replication mode is transferred into the
female (Fig. 8.19). Immediately upon entry of the single strand into the
female, the complementary strand is synthesized. If no breakage occurs during
transfer, all the F-factor DNA, including the portion initially left behind at the
break, can be transferred into the female cell. The F-factor thus codes for
transfer into female cells of itself and also any DNA to which it is connected.
If the F-factor is integrated within the chromosome, then its mobilization in
conjugation transfers the chromosome as well as itself. Cells which transfer
their chromoso - mal genes to recipient cells are called Hfr for high frequency
of recom-bination, whereas the cells which contain F-factors separate from the
chromosome are called F or F’, and the females are called F-.
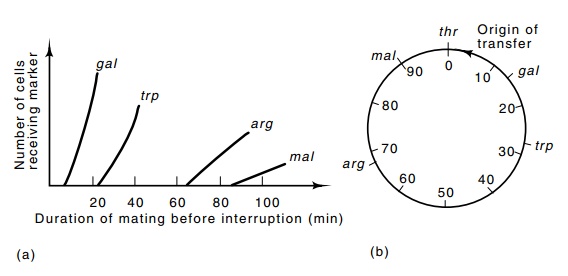
A little over 100 minutes are required for transfer of the entire Escherichia coli chromosome to a female. Genes located anywhere onthe chromosome can be mapped by determining the timing of their transfer
to female cells. To do this mating is initiated by mixing male and female
cells. Genetic markers on one side of the F-factor will be transferred to the
female cells with only a few minutes of mating, while markers on the opposite
side of the chromosome will not be transferred for over half an hour.
Random breakage of the DNA during mating from an Hfr to an F-often interrupts the transfer so that only infrequently can the complete chromosome be transferred. This is a blessing in disguise however, for in addition to the time of transfer, the frequency of a marker’s transfer also indicates its chromosomal position with respect to the integrated F-factor. More than one integration site of F has been found on the chromosome, so that a variety of origins of transfer are available and the entire chromosome can be easily mapped. Once mating has been started and been allowed to proceed for a period of time, any further transfer of markers can be stopped merely by vigorously shaking the culture to separate the mating couples.
A mating
experiment might be performed in the following way. Male Arg+ Gal+
Leu+ Sms (streptomycin-sensitive) and female Arg-
Gal- Leu-Smr cells would be grown to densities
of about 1 × 108
per ml and mixed together in equal portions. The cells would then be shaken
very gently. At intervals, a sample of cells would be taken, vigorously shaken
so as to separate mating couples and dilutions spread on
streptomycin-con-taining medium so as to select for Arg+ or Gal+
or Leu+ Smr recombi-nants (Fig. 8.20).
F-factors
facilitate genetic study with Escherichia
coli in a second way. They need not be associated with the chromosome.
Instead, they can be autonomous DNA elements existing alongside the chromosome.
The transfer properties remain the same for an F-factor in this position, but
the genetics are slightly altered in two ways. First, the extra-chro-mosomal
F-factor can have some chromosomal genes associated with it, in which case it
is called an F’-factor. Presumably these genes were
Related Topics