Chapter: Genetics and Molecular Biology: Genetics
Heteroduplexes and Genetic Recombination
Heteroduplexes and Genetic Recombination
Having
considered the existence and use of genetic recombination, we are ready to
consider how it comes about. Genetic recombination yields a precise cut and
splice between two DNA molecules. Even if one DNA

molecule
were to have been cut, it is hard to imagine how an enzyme could know where to
cut the other DNA duplex so as to produce the perfect splices that genetics
experiments show occur. The difficulty can be largely overcome by a mechanism
utilizing the self-complementary double-stranded structure of DNA. A denatured
portion of one duplex could anneal to a denatured portion of complementary
sequence from the other duplex (Fig. 8.12). This would hold the two DNA molecules
in register while the remainder of the recombination reaction proceeded.
The life
cycle of yeast permits a direct test of the model outlined above. A diploid
yeast cell undergoes recombination during meiosis, and the two meiotic cell
divisions yield four haploid spores. These four spores can be isolated from one
another and each can be grown into a colony or culture. In essence, the cells
of each colony are identical copies of each of the original recombinants, and
the cells can be tested to determine the genetic structure of the original
recombinants. If one of a pair of homologous chromosomes contains a mutation
and the other does not, generally two of the four resulting spores will contain
the mutation and two will not.
Consider
the situation resulting from melting portions of the du-plexes and base pairing
between complementary strands of two homolo-gous yeast chromosomes in the
process of genetic recombination. The region of pairing may include the
mutation. Then a heteroduplex forms that contains the mutant sequence on one
strand and the wild-type sequence on the other (Fig. 8.13). As discussed,
mispaired bases are subject to mismatch repair and, if it occurs, the yeast
repair system in this case has no apparent reason to choose one strand to repair
in preference to the other. Therefore strands may be correctly or incorrectly
repaired, so the final outcome could be three copies of the wild-type or mutant
sequence and one copy of the other in the meiosis from a single yeast cell. In
total, a single yeast cell can produce one or three progeny spores containing
the marker from one of the original chromosomes. This phenomenon is called gene
conversion. It is experi-mentally observed and consequently it is reasonable to
expect that pairing between complementary strands of recombinant partners
oc-curs during recombination. Without heteroduplex formation and mis-match
repair, there is no easy way to generate any ratio other than 2:2.
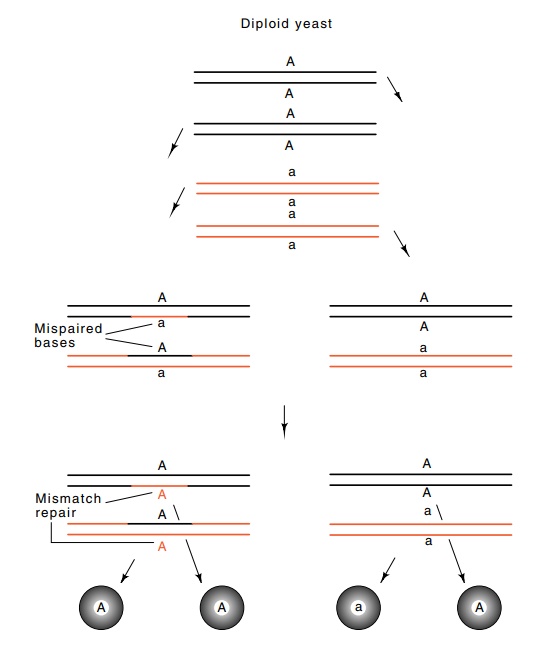
Figure
8.13 Gene conversion in yeast. A
diploid (A/a) undergoes meiosis,which produces heteroduplexes A-a that are both
repaired to A-A.
Related Topics